Nonstructural carbohydrates influence water relations in Pinus ponderosa

Non‐structural carbohydrates influence water relations in Pinus ponderosa Aurora Bayless The Sala lab of Plant Physiological Ecology Division of Biological Sciences University of Montana

Why study water relations in Ponderosa pine? • Drought is predicted to increase with climate change • Drought stress mortality is a major driving factor in forest ecology 2, 562 matches Mainsfield search 123, 000 articles google scholar • Forests hold lots of carbon, and influence global cycles • The specific mechanisms behind drought related mortality are still unknown Shinneman et al. 2016
 Sugars are hydrophilic Structural Carbohydrates (cellulose)")
Photosynthesis makes carbohydrates Non‐Structural Carbohydrates (NSC) Sugars are hydrophilic Structural Carbohydrates (cellulose)

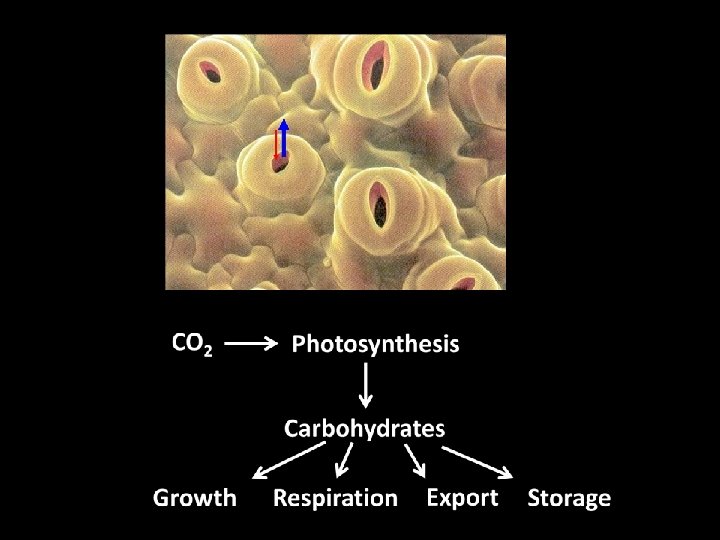
Making sugar requires water loss. Water loss increases xylem tension.

Xylem tension is dangerous! Plants must prevent water loss or the water column will break leading to catastrophic hydraulic failure

Plants can close stomata to prevent water loss However, closing stomata to prevent water loss prevents photosynthesis carbon starvation
 hydraulic failure,")
Two main hypotheses for the mechanisms of drought stress mortality: • 1) hydraulic failure, mortality is caused by xylem cavitation • We know this • 2) Carbon starvation happens as a result of stomatal closure (Mc. Dowell, et al. 2008). • We don’t know the threshold yet

NSC is not used as expected drought control Drought does not deplete starch Galvez et al. 2011. Tree Physiol. Sala & Hoch 2009. PCE Wiley et al. 2015. Tree Physiol. Palacio et al. 2008. Funct. Ecol.

Why so much NSC? Metabolism Reservoir Osmotic role NSC may help retain and move water

• It is critical to know how NSC affects hydraulic function to predict mortality. • First we need to know if there is a threshold for carbon starvation • Then, we can ask how carbon starvation may drive the hydraulic failure to critical thresholds and vice versa
 Non‐Structural Carbohydrates are related to hydraulic function. • 2) NSC")
Research hypotheses: • 1) Non‐Structural Carbohydrates are related to hydraulic function. • 2) NSC below a certain threshold causes mortality.

7 CONTROL TREATMENT
 Stems Needles 4")
Non‐Structural Carbohydrates decreased in the dark 8 Roots 6 NSC (%) Stems Needles 4 2 0 CI 1 2 3 # weeks in dark 4 Stepwise NSC decline with dark exposure 5 6
 8 6 4")
Non Structural Carbohydrates decreased in the dark Whole Plant NSC (%) 8 6 4 2 0 CI 1 2 3 # weeks in dark 4 Stepwise NSC decline with dark exposure. Weeks 1‐ 3 and weeks 4‐ 6 have similar NSC 5 6

Hypothesis 1 • Non‐Structural Carbohydrates are related to hydraulic function.

Pre‐Dawn Water potential
 Decreases with dark exposure despite irrigation ‐ 1 Y (MPa) Increasing")
Water Potential (Y) Decreases with dark exposure despite irrigation ‐ 1 Y (MPa) Increasing water stress 0 ‐ 2 ‐ 3 ‐ 4 CI 1 2 3 # weeks in dark 4 5 6

High water stress despite irrigation is related to declining NSC 1 3 2 4 5 6 C

Water stress occurs at about 40% NSC relative to controls 1 3 2 4 5 6 C

Positive carbon assimilation occurs after one and four weeks of dark exposure 6 A (mmol m‐ 2 s‐ 1) 4 2 0 ‐ 2 ‐ 4 CI 1 2 3 4 5 6 # weeks in dark Declining NSC after four weeks of dark exposure stimulates C assimilation. This could be a compensation mechanism to make up for lost carbohydrates, because of shade.

Increasing water stress Higher water stress despite irrigation related to declining NSC

Increasing water stress Higher water stress despite irrigation related to declining NSC‐depleted plants with low water stress tend to maintain positive photosynthesis. This indicates that Carbon is needed to retain water.
 NSC below a certain threshold causes mortality.")
Hypothesis 2 • 2) NSC below a certain threshold causes mortality.

In well watered groups the first mortality occurs after three weeks of dark exposure 90 80 70 % Mortality 60 50 40 30 20 10 0 0 1 2 Weeks of Dark 3 4 5

Below 40% NSC relative to controls plants are unable to maintain hydraulic function and mortality increases

What does this mean for the future? • Drought is a major issue, and we need to model it to understand mortality and Carbon cycles. • How is this research important for land managers, modelers, forest managers, et cetera? • We need to start considering the interactions of hydraulic function and Non Structural Carbohydrates in order to model future tree mortality.

Works cited • • • Allen, C. D. , A. Macalady, H. Chenchouni, and et al. "A global overview of drought and heat‐induced tree mortality reveals emerging climate change risks forests. " Forest Ecology and Management, 2010: 660– 684. Bonan, Gordon B. "Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. " Science Vol. 320 no. 5882 , 2008 : 1444‐ 1449. DNRC, Forestry Assistance Bureau of Forestry Division of the. Montana Conservation Seedling Nursery. January 1, 2015. http: //www. dnrc. mt. gov/forestry/Nursery/default. asp (accessed January 1, 2015). Galvez, David A. , S. M. Landhäusser, and M. T. Tyree. "Root carbon reserve dynamics in aspen seedlings: does simulated drought induce reserve limitation? . " Tree Physiology 31. 3 (2011): 250‐ 257. Hoch, G. , M. Popp, and Ch. Korner. "Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline. " Oikos, Vol. 98, 2002: 361‐ 374. Mcdonald, P. M. , C. N. Skinner, and G. O. Fiddler. "Ponderosa pine needle length: and early indicator of release treatment effectiveness. " Canadian Journal of Forestry Research Vol. 22, 1992: 761‐ 764. Mc. Dowell, N. G. , and S. Sevanto. "The mechanisms of carbon starvation: how, when, or does it even occur at all? " New Phytologist, 2010: 264– 266. Mc. Dowell, Nate G. , David J. Beerling, David D. Breshears, and Rosie A. Fisher. "The interdependence of mechanisms underlying climate‐driven vegetation mortality. " Trends in Ecology and Evolution, 2011: 1‐ 10. Mc. Dowell, Nate, et al. "Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? . " New Phytologist, 2008: 719‐ 733. Mc. Dowell, Nathan. "Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. " Plant Physiology, 2011: 1051‐ 1059. Mc. Millin, J. D. , and M. R. Wagner. "Effects of water stress on biomass partitioning of ponderosa pine seedlings during primary root growth and shoot growth periods. " Forest Science, Vol 41, No 3, 1995: 594‐ 610. O’Brien, Michael J. , David F. R. P. Burslem, Alexa Caduff, John Tay, and Andy Hector. "Contrasting nonstructural carbohydrate dynamics of tropical tree seedlings under water deficit and variability. " New Phytologist, 2014: 1‐ 12. • Palacio, Sara, et al. "Browsed Betula pubescens trees are not carbon‐limited. " Functional Ecology 22. 5 (2008): 808‐ 815. • Repetto, G. , A. del Peso, and J. L. Zurita. "Neutral red uptake assay for the estimation of cell viability/cytotoxicity. " Nature Protocols, 2008: 1125‐ 1131. • Sala, Anna, and Günter Hoch. "Height‐related growth declines in ponderosa pine are not due to carbon limitation. " Plant, Cell & Environment 32. 1 (2009): 22‐ 30. • • Sala, A. , D. R. Woodruff, and F. C. Meinzer. "Carbon dynamics in trees: feast or famine? " Tree Physiology, 2011: 1‐ 12. Sala, A. , F. Piper, and G. Hoch. "Physiological mechanisms of drough tinduced tree mortality are far from being resolved. " New Phytologist, 2010: 274– 281. Sala, Anna, and Robert Nowak. "Ecophysological responses of three riparian graminoids to changes in the soil water table. " Biological Faculty Publications University of Montana, 1997: 835‐ 843. Solomon, Suzan, Gian‐Kasper Plattner, Reto Knutti, and Pierre Friedlingstein. "Irreversible climate change due to carbon dioxide emissions. " PNAS vol. 106 no. 6, 2008: 1704– 1709. Trenberth, Kevin E. "Changes in precipitation with climate change. " Inter‐Research Climate Research vol. 47 no. 1‐ 2, 2011: 123‐ 138. Turner. "Measurement of plant water status by the pressure chamber technique. " Irrigation Science 9 (1988): 289‐ 308. van Mantgem, P. J. , N. L. Stephenson, et al. , and J. C. Byrne. "Widespread increase of tree mortality rates in the Western United States. " Science, 2009: 521– 524. Wennerberg, Sarah. "Ponderosa Pine Pinus ponderosa P. & C. Lawson. " USDA NRCS Plant Guide, 2004: 1‐ 4. • Wiley, Erin, and Brent Helliker. "A re‐evaluation of carbon storage in trees lends greater support for carbon limitation to growth. " New Phytologist 195. 2 (2012): 285‐ 289. • • • image credits: http: //ehinger. nu/ Buzzle. com https: //online. science. psu. edu/ http: //www. viewzone 2. com/hifruc. htmlhttp: //www. biosciguy. com/notes%20 on%20 water%20 potential. htm Shinneman, D. J. , Means, R. E. , Potter, K. M. , Hipkins, V. D. , 2016, Exploring climate niches of ponderosa pine (Pinus ponderosa Douglas ex Lawson) haplotypes in the western United States‐ implications for evolutionary history and conservation: PLOS ONE, v. 11, no. 3, p. e 0151811, http: //dx. doi. org/10. 1371/journal. pone. 0151811.
 • Anna")
Thank you! Sala Lab Physiological Plant Ecology • Sean Pinnell (MS Student) • Anna Sala (PI) • Beth Roskilly (MS student) • Gerard Sapés (Ph. D Student)

QUESTIONS?
- Slides: 30