MUTANTI WOX geni WOX Cosi chiamati perch contengono










 WT ton Nei mutanti ton (fass) Le piante sono deformate a")














- Slides: 29
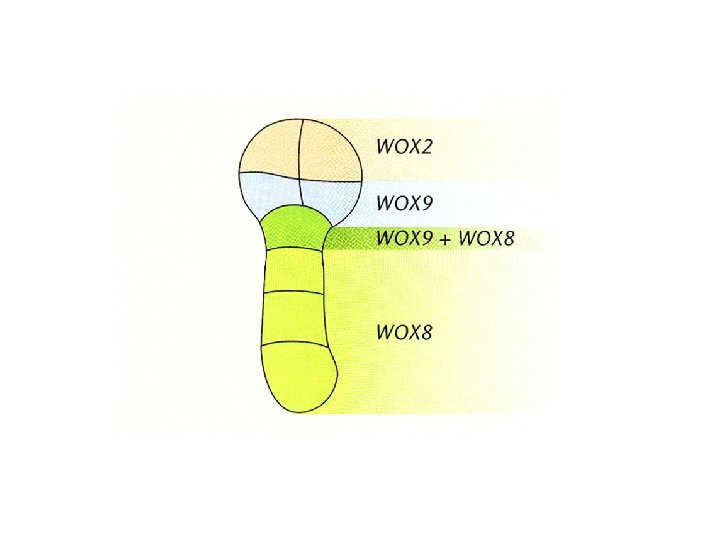
MUTANTI WOX geni WOX Cosi chiamati perché contengono il motivo di binding al DNA dei geni HOMEOBOX e per la loro somiglianza a WUSCHEL Ruolo nella formazione del pattern assiale Nella prima divisione dello zigote WOX 2 si accumula nella cellula apicale, WOX 8 e WOX 9 in quella basale Mutanti wox 2 mostrano divisione cellulare alterata nel proembrione WT wox 2 -1 wox 2 -2
Alllo stadio di 8 cellule i differenti domini dell’embrione sono distinti da un diverso e complementare profilo di espressione dei geni WOX
Espressione dei geni WOX in stadi precoci dell’embriogenesi
Mutanti wox 1, 2, 5, 8, 9 isolati da una collezione di mutanti d’inserzione wox 2 difetti nello sviluppo della porzione apicale wox 9 ridotto numero di cellule nella parte basale dell’embrione (ipocotili ridotti) wox 2: specificazione del destino della cellula apicale (porzione superiore dell’embrione) wox 9 specificazione delle cellule basali dell’embrione (ipocotile e radice) wox 1, wox 3, wox 5, wox 8: assenza di fenotipo Ridondanza?
Mutazioni nei geni MONOPTEROS e BODENLOS alterano il pattern di espressione di WOX 9 (inibito il passaggio dall’ipofisi alla parte inferiore dell’embrione) Gradienti di auxina regolano l’espressione dei geni WOX? WOX bersaglio di Monopteros?
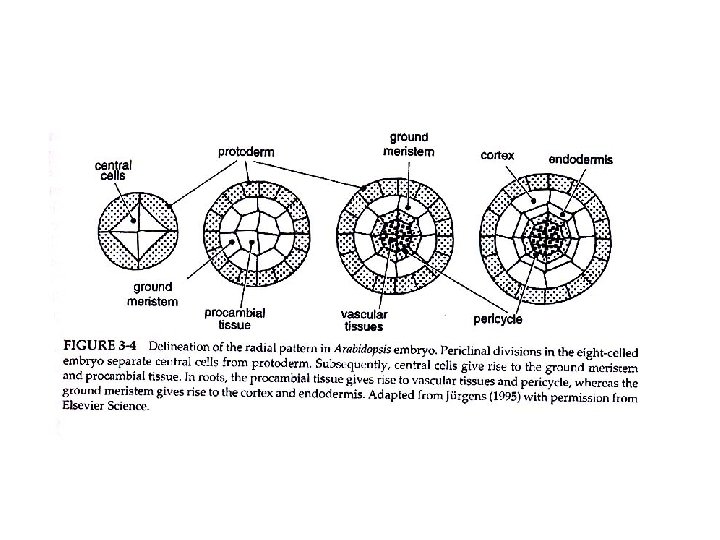
FORMAZIONE DEL PATTERN RADIALE
Allo stadio globulare divisioni periclinali dividono l’embrione in tre regioni radialmente distinte: Cellule più esterne : strato singolo di PROTODERMA si differenzia in EPIDERMIDE Cellule sotto il protoderma: MERISTEMA FONDAMENTALE origina il CORTEX e l’ENDODERMIDE Cellule più interne: PROCAMBIO genera i TESSUTI VASCOLARI (e nella radice il PERICICLO)
ANTICLINALI: divisioni perpendicolari alla superfice dell’organo PERICLINALI: divisioni parallele alla superficie dell’organo
Sequenza di formazione del pattern radiale
Come per il pattern assiale nella formazione del Pattern Radiale non sembra indspensabile una precisa sequenza di divisioni cellulari più rilevante l’informazione posizionale MUTANTE TONNEAU 2 (TON) (FASS)
TON 2 (FASS) WT ton Nei mutanti ton (fass) Le piante sono deformate a causa di divisioni cellulari in eccesso e piani di divisione alterati; tuttavia formano tessuti riconoscibili e nelle posizioni corrette sezione di radice TON 2: Protein fosfatasi che regola le funzioni del citoscheletro; non si forma la banda preprofasica di microtubuli; piani di divisione ed espansione irregolari
Le divisioni cellulari sono anormali ma i tessuti si formano nelle posizioni corrette (Il solo parenchima corticale è pluristratificato nel mutante) Il differenziamento non dipende dalla divisone cellulare di per sé
Formazione del pattern radiale Basi molecolari meno chiare Probabilmente coinvolta informazione posizionale ma meccanismi non noti quale morfogeno?
DIFFERENZIAMENTO DEL PROTODERMA posizione unica, strato superficiale, non circondato da tutti i lati da altre cellule Identificati due geni implicati nella determinazione della identità delle cellule del protoderma (foglie) ATML 1 PDF 2 (arabidopsis thaliana meristem layer 1) (protodermal factor 2) Espressi precocemente durante l’embriogenesi solo nello strato superficiale di cellule Codificano entrambi per fattori di trascrizione HOMEODOMAIN e sono espressi negli stadi iniziali dell’embriogenesi nelle cellule dello strato più esterno dell’embrione (riconoscono una sequenza sul promotore di molti geni epidermide specifici)
Mutazione atml 1/pdf 2 Il mutante ha epidermide con caratteristiche di mesofillo
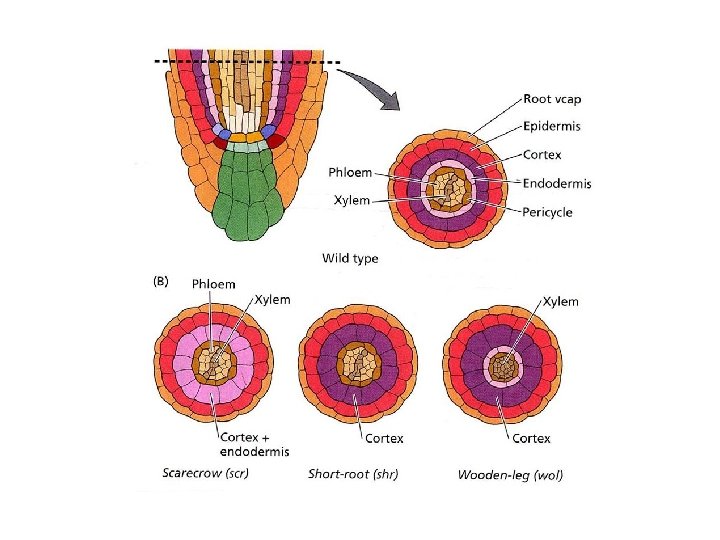
DIFFERENZIAMENTO DI CORTEX/ENDODERMIDE Nella radice e nell’ipocotile durante l’embriogenesi Richiesta l’espressione dei geni SHORT ROOT (SHR) SCARECROW (SCR) Isolati da mutanti di Arabidopsis con radici corte In entrambi il cortex non è differenziato dall’endoderma: üsingolo tipo cellulare a caratteristiche miste (scr) üsolo cellule corticali (shr)
SCR wt E C En scr 1 wt scr
Mutazione shr: singolo strato di Cortex SHR necessario per specificare l’identità delle cellule dell’endodermide
shr wt shr Marcatori del cortex Ab anti epitopo di parete di cortex e epidermide (lo strato mutato ha caratteristiche di cortex) AX 92: : GUS promotore per cortex
scr : espressione di entrambi i caratteri di endodermide e cortex ma incapacità di separarli in due strati distinti
L’espressione del m. RNA di SHR è ristretta nei tessuti provascolari
espressione di SCR e SHR in embrioni di Arabidopsis WT e shr (microscopia confocale con GFP sotto il controllo dei promotori di SCR o SHR. SCR: : GFP; SHR: : GFP Wt SHR E C En Tessuto provascolare Wt shr c/ei qc SCR Strato di cellule mutate En c/ei qc c/e i= iniziali cortex/endodermide
La proteina SHR espressa nella stele è in grado di migrare nell’endodermide (m. RNA di SHR) promotore-SHR: : GFP (proteina SHR) SHR: GFP (proteina chimerica)
ØIl prodotto del gene SHR viene espresso nella stele e poi trasportato nello strato adiacente esterno dove induce i tratti di endodermide (cell to cell signaling) ØIl prodotto del gene SCR è espresso nel centro quiescente, nelle iniziali del cortex/endodermide e nell’endodermide ØSHR è necessario alla espressione di SCR
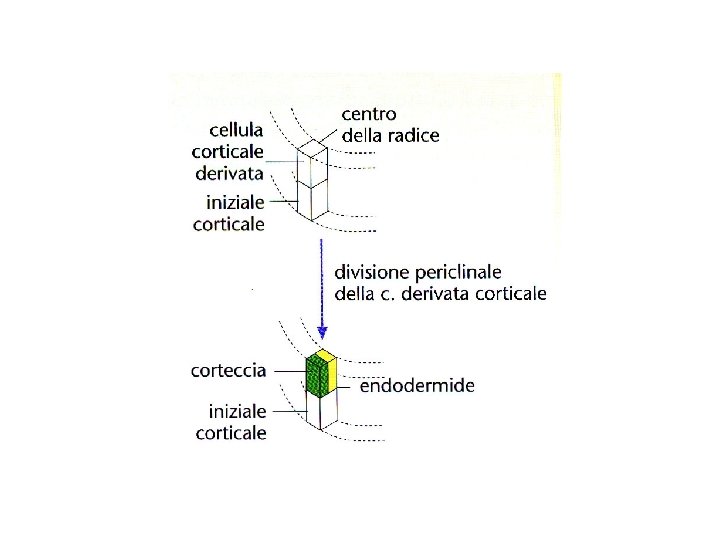
Derivata Iniziale corticale