Molekulrn genetika I Molekulrn zklad ddinosti vod molekulrn



 RNA = ribonukleová kyselina (ribonucleic acid)")








 - přenos aktivovaných AA na místo biosyntézy")










 •")






 Strukturní gen - úsek DNA, který je přepisován do primární struktury polypeptidu:")


 (polymerase chain reaction): - r. 1986")






 - dříve: klasické metody - při testování parentity šlo pouze o")








")
")

- Slides: 55
Molekulární genetika I. Molekulární základ dědičnosti
Úvod • molekulární genetika - studium procesů podmiňujících dědičnost a proměnlivost na molekulární úrovni • genetická informace (GI) je zakódována v DNA - uložena v J buňky v chromozómech • odhalení genetického kódu (GK) - Watson a Crick (1953) - zlomový objev pro další rozvoj molekulární genetiky • v posledních desetiletích bouřlivý rozvoj MG - projekty: HUGO (Human Genome) - mapování lidského genomu Bov. Map, Pig. Map, Chick. Map
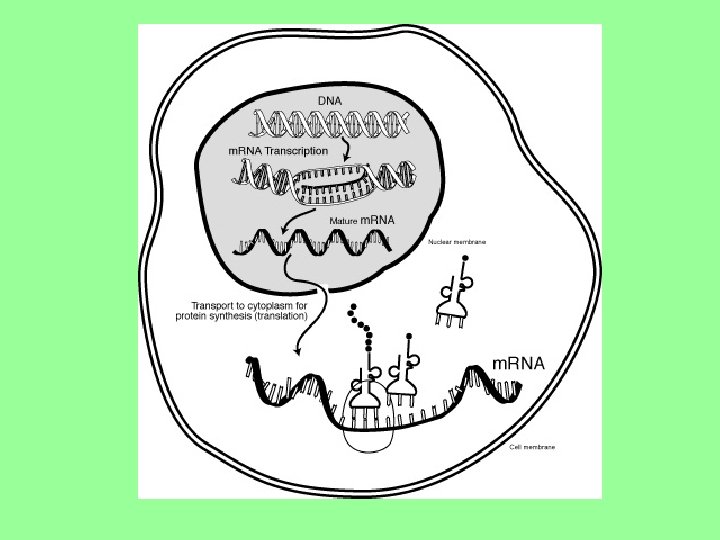
Tok genetické informace Exprese genetické informace je komplexní proces a lze si jej představit jako informační tok (přesun informace pomocí informačních molekul z sekvence nukleotidů do sekvence AA) v buňce. Souhrnně tyto procesy (směr "toku" genetické informace) představují tzv. Centrální dogma molekulární genetiky: –> protein“ “DNA –> RNA
Nukleové kyseliny DNA = deoxyribonukleová k. (deoxyribonucleic acid) RNA = ribonukleová kyselina (ribonucleic acid) dvouvláknová - dvoušroubovice jednovláknová Funkce: DNA - nositel GI *) RNA - podíl na realizaci GI *) u některých virů je nositelem GI: RNA - např. medicínsky důležité viry: - virus chřipky - virus HIV (Humane Immunodeficiency Virus) - vyvolává AIDS chromozómy se skládají z řady makromolekul: - cca 60– 90 % tvoří zhruba ve stejném poměru DNA a histony (bázické proteiny), - dále nehistonové proteiny a molekuly RNA
DNA
Struktura NK Základní stavební kamen NK: nukleotid = cukr + fosforečný zbytek + dusíkatá báze - pětiuhlíkatá ribóza (v RNA) - deoxyribóza (v DNA) na 3. a 5. atomu uhlíku fosfátová skupina spojuje molekuly cukru navzájem do řetězce na 1. atomu uhlíku cukru nukleotid X nukleosid - purinové A, G - pyrimidinové báze T, C - DNA: A, T, G, C pětiuhlíkatý cukr - RNA: A, U, G, C + fosfátová skupina + dusíkatá báze + deoxyribóza
Cukerná složka u RNA u DNA Zbytek H 3 PO 4
Dusíkaté báze Purinové adenin A guanin G Pyrimidinové thymin T cytosin C uracil U
Nukleotid
Řetězec nukleotidů 3´- 5´ V DNA - dva protisměrné nukleotidové řetězce: 1. ve směru 5´- 3´ „dolů“ („po směru“) 2. 3´- 5´ „nahoru“ („proti směru“) navzájem jsou spojeny do dvoušroubovice prostřednictvím vodíkových můstků mezi komplementárními bázemi A - T (2 můstky) a G - C (3 můstky).
DNA Strukturu DNA můžeme přirovnat k točitým schodům, jejichž "bočnice" jsou tvořeny spojením cukr - fosfátová skupina (monotónně se opakuje). Poloviny jednotlivých stupňů jsou tvořeny dusíkatou bází, která se napojuje na druhou polovinu - komplementárního partnera ze druhého vlákna.
Komplementarita bází A - T DNA A - U RNA C - G C - G Příklad komplementarity: 1. vlákno 2. vlákno A G T C T CA G
RNA Typy RNA: 1. transferová (t. RNA) - přenos aktivovaných AA na místo biosyntézy bílkovin ( ribozómy) 2. ribosomální (r. RNA) - součást ribozómů, které jsou místem biosyntézy bílkovin 3. mediátorová (m. RNA) - přenos GI z DNA na bílkoviny (messenger = poslíček) (ve formě specifické sekvence nukleotidů) - různá délka (několik set až tisíc nukleotidů)
t-RNA Transferová RNA - tvar připomínající trojlístek
r. RNA m. RNA
Rozdíly mezi DNA a RNA DNA • deoxyribóza • A, T, G, C • dvouvláknová • 1 typ DNA *) • stejná fce **) cukr báze počet vláken počet typů funkce *) u všech živých organismů **) uchovávání a přenos GI RNA • ribóza • A, U, G, C • ± 1 vláknová • 3 typy RNA • rozličné fce
Přenos GI DNA RNA Protein
Replikace, transkripce, translace
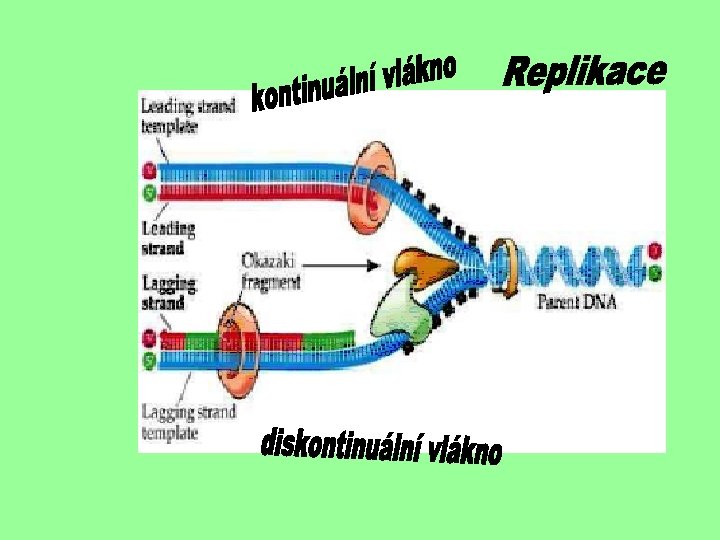
Replikace DNA • • Struktura ve formě dvoušroubovice a komplementarita bází umožňují semikonzervativní replikaci DNA - z 1 mateřské dvoušroubovice DNA → 2 dceřinné (každá z nich 1 vlákno původní - mateřské a 1 nově syntetizované). Replikace je zajišťována DNA polymerázami. Replikační vidlička - místo, kde probíhá syntéza nového vlákna Dvoušroubovice DNA se rozpojí, každé vlákno je předlohou pro vznik komplementárního vlákna DNA. Postupně jsou na nově vznikající vlákno připojovány nukleotidy, které mají dusíkatou bázi komplementární k bázi na původním vláknu DNA. Syntéza probíhá v obou směrech, obě replikační vidličky se od sebe vzdalují a vytvářejí tak útvar, který je možné pozorovat v elektronovém mikroskopu jako replikační bublinu. V prokaryontních buňkách je DNA syntetizována jako jednotka, tj. celý chromozóm je jeden replikon, v eukaryontních je rozčleněna do více částí - replikonů. Replikace probíhá v S fázi buněčného cyklu.
Schéma replikace
Replikon
Transkripce • podle předlohy v DNA syntetizována RNA • 1 mechanismus transkripce pro všechny RNA • Přepisována, transkribována je sekvence nukleotidů v DNA, podle které vzniká komplementární sekvence nukleotidů v RNA. • Tento proces je enzymaticky zajišťován RNA polymerázami. • Signální sekvence DNA před genem, na které se váže RNA polymeráza se nazývá promotor. Na promotor se naváží transkripční faktory (proteiny), dále RNA polymeráza a za přítomnosti volných ribonukleotidů probíhá syntéza nového řetězce m. RNA. • Produktem transkripce je prekurzorová pre-m. RNA (kromě vlastního genu obsahuje také introny a okrajové části genu) → maturace (dozrávání) m. RNA - dojde k enzymatickému rozštěpení pre-m. RNA a spojení exonů (kódujících částí). Introny (nekódující části) zůstávají v jádře. Tento proces se nazývá splicing (=sestřih).
Schéma transkripce
Posttranskripční úpravy • po ukončení transkripce - vznik pre-m. RNA => zrání (maturace) • pre-m. RNA - 2 typy úseků: exony a introny • SPLICING = sestřih - „vystřižení“ intronů Schéma splicingu
Translace • = přepis informace o sekvenci nukleotidů v m. RNA do sekvence AA (aminokyselin) ve vznikajícím proteinu (překlad nukleotidů do pořadí AA v polypeptidovém řetězci) • probíhá na ribozómech (komplex několika typů r. RNA a proteinů) • volné AA jsou vázány na t. RNA, transportovány do ribozómu, kde t. RNA nasedá svým antikodonem (specifická trojice bází) na kodon na m. RNA • pořadím tří bází na kodonu je dána AA, která bude na příslušném místě v syntetizovaném proteinu • na sousední kodon nasedá další t. RNA s AA, mezi aminokyselinami vzniká peptidová vazba, dále ve směru 5´- 3´ nasedají další t. RNA a navazují se další AA
Schéma translace
Translace
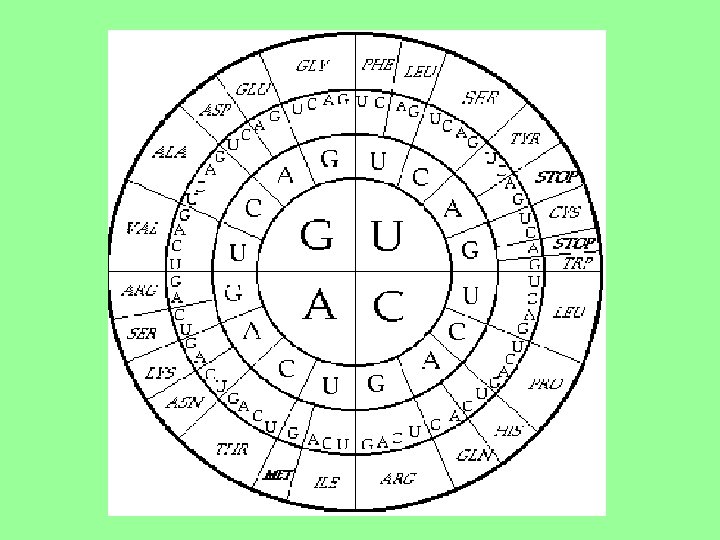
Genetický kód - soubor pravidel, podle kterých se genetická informace uložená v DNA, resp. RNA, převádí na primární strukturu bílkovin - tj. pořadí aminokyselin v řetězci Vlastnosti - GK je: 1. tripletový - pořadí tří nukleotidů určuje aminokyselinu - trojice bází kóduje 1 AA (trojice nukleotidů = kodon, triplet) 2. univerzální - u všech organismů kóduje 1 a ten samý triplet stejnou AA (pravidlo platí pro velkou většinu kodonů, u některých organismů mají některé kodony odlišný smysl) 3. degenerovaný (nadbytečný) - 1 AA může být determinována více kodony (1 AA je kódována více triplety) 4. bez mezer a nepřekrývající se - jednotlivé kodony v molekule NK následují bezprostředně za sebou
Ze tří různých bází v kodonu je možné sestavit celkem 64 kombinací !
Gen = úsek DNA, která má: 1. specifickou funkci v buňce a v celém organismu - musí být schopen utvářet dědičný znak nebo spolupracovat při utváření několika takovýchto dědičných znaků 2. je schopen vytvářet přesné vlastní kopie - přenášet svou funkci z generace na generaci 3. charakteristickou vlastností genu je schopnost náhle změnit svou strukturu a tím i funkci, tj. podléhat mutacím. Zmutovaný gen replikuje svou změněnou podobu. Lokus - místo na chromozomu, na kterém je gen umístěn Alely - odlišné formy téhož genu, které u různých jedinců téhož druhu alterují na jednom lokusu (uspořádání lokusů je pro druh konstantní)
Gen 1) Strukturní gen - úsek DNA, který je přepisován do primární struktury polypeptidu: Strukturní gen - transkripce - m. RNA - translace - polypeptid. 2) Gen pro funkční RNA - úsek DNA, který je přepisován do t. RNA, r. RNA nebo jiného typu RNA, avšak neprobíhá podle něj translace. 3) Gen jako regulační oblast - úsek DNA, identifikovaný specifickým proteinem, který zahajuje nebo zastavuje transkripci. Podle regulační oblasti se tedy netvoří žádný produkt, ani RNA, ani polypeptid. Struktura genomu U eukaryot je DNA: v jádře buňky v chromozomech, v mitochondriích (živočiš. buňky), v chloroplastech (rostlinné buňky). Mimojaderné geny však hrají v eukaryontní buňce významnou roli, u rostlin probíhá v chloroplastech fotosyntéza, u živočichů v mitochondriích oxidázový cyklus.
II. Molekulárně genetické metody - úvod
Využití MG metod Na úrovni lidské DNA: - identifikace sekvence podmiňující vznik dědič. onemocnění - určování genetické identity jedinců - určování parentity (rodičovství) - průkaz původu biol. materiálu v soudním lékařství apod. Na úrovni DNA nebo RNA bakteriálních nebo virových infekčních agens: - rychlá detekce - využití v klinických vzorcích - nahrazení zdlouhavého testování původce onemocnění na základě až vytvořených protilátek (ELISA test)
Metoda PCR - metoda polymerázové řetězové reakce (PCR) (polymerase chain reaction): - r. 1986 Saiki a Mullis - Nobelova cena - umožňuje namnožit DNA nebo její fragmenty do množství potřebného pro diagnostické účely
Principy molekulárně genetické diagnostiky Izolace nukleových kyselin - základem odstranění proteinů, které tvoří s DNA komplex (fenol-chloroformová extrakce, vysolení, působnost různých enzymů apod. ) Diagnostika NK / fragmentů NK - podstata: rozštěpení DNA na fragmenty (restrikčními enzymy) a jejich následná identifikace (např. s využitím Southernova přenosu, PCR a jejích modifikací apod. )
Molekulárně genetická diagnostika dědičných chorob Lidský genom - asi 30 000 genů - každý gen je potenciálně polymorfní - výskyt v různých variantách (alelách) vzniklých mutačním procesem - řada z těchto mutací může vést ke vzniku danou mutací podmíněné choroby Princip přímé a nepřímé MG diagnostiky dědičných chorob: poznání rozdílu v DNA původní a mutované alely a identifikace těchto rozdílů
Přímá diagnostika - detekce mutací - metody umožňují identifikovat typ bodové mutace u jejího nositele (podle obrazu na elektroforeogramu) - např. Duchenneova svalová dystrofie
Detekce deleční formy Duchenneové svalové dystrofie Fragmenty 3, 3, resp. 3, 1 kb jsou součástí X-chromozomového genu, který je za toto onemocnění zodpovědný. Postižený syn: 0, 8 kb - není přítomný ani u jednoho rodiče, vznikl jako důsledek vnitrogenové delece, která zkrátila restrikční fragment z 3, 1 na 0, 8 kb. (Ferák a Sršeň 1990)
Nepřímá diagnostika - v případech, kdy u pacienta nelze přímo zjišťovat přítomnost mutace, opírá se molekulárně genetická diagnostika o vyšetření polymorfismu charakterizovaném chyběním nebo přítomností cílového místa pro určitý restrikční enzym - nejčastější metodou: metoda polymorfismu délky restrikčních fragmentů - RFLP (restriction fragment lenght polymorphism) - např. diagnostika hemofilie A
Výsledek elektroforetické separace restrikč. fragmentů
Testování parentity (rodičovství) - dříve: klasické metody - při testování parentity šlo pouze o vyloučení nesprávně uvedeného rodiče, nikoliv o jeho potvrzení - dnes: aplikace MG metod - spolehlivější, výsledkem může být takřka 100% potvrzení rodičovství - využití vysoce polymorfních mikrosatelitních lokusů - např. D 1 S 7 s teoreticky očekávanými 2400 alelami alternujícími na tomto lokusu - > 99% jedinců je na tomto lokusu heterozygotních => shodné genotypy např. více potenciálních otců mají jen nízkou pravděpodobnost incidence
Molekulárně genetický důkaz biolog. materiálů a infekčních agens Rozšíření využití PCR —> rychlost, extrémní citlivost - klinická mikrobiologie - virologie - hodnocení kvality potravin - vychází se z průkazu patogenu na základě identifikace jeho druhově specifické sekvence nukleotidů v DNA - pro průkaz patogenu postačí teoreticky 1 molekula DNA - obdobné metodiky jsou používány pro rozlišení např. biologických stop krve, sperma, slin, vlasů, epitelií a jiných biologických materiálů v soudním lékařství nebo kriminalistice
Druhově specifická amplifikace sekvence DNA pro různé druhy hosp. zvířat a analyzovaný vzorek => identifikovaný vzorek - směs dvou mas
Postup při realizaci PCR-RFLP
PCR - probíhá inkubací vzorků při třech teplotách odpovídajících třem krokům v amplifikačním cyklu: denaturaci, annealing a elongaci
RFLP - pomocí RFLP se identifikují alely na základě přítomnosti nebo absence specifického restrikčního místa - namnožená genomová DNA (PCR produkt) je štěpena příslušnou restrikční endonukleázou a separována pomocí elektroforézy na agarózovém gelu - vizualizace DNA se provádí pomocí ethidiumbromidu - reakční směs pro štěpení PCR produktu: pufr pro restrikční endonukleázu, enzym restrikční endonukleáza a příslušné množství destilované H 2 O, do směsi se přidávají vzorky DNA - teplota inkubace: 37 °C - enzymatické štěpení specifických míst sekvence DNA, které rozpoznává daný enzym - po inkubaci se do jamek v gelu nanese pipetou vzorek - vyhodnotí se fragmenty vzorků podle markeru, tvořící žebříček fragmentů
Determinace pohlaví pomocí PCR – vzorek kostí z pohřebiště z 8. -9. stol. (Kolárik a kol. , 2002)
Hemofilie
Huntingtonova chorea
Genetický polymorfismus LEP genu prasat pomocí PCR-RFLP (Bábelová a kol. , 2002)
Genetický polymorfismus ESR genu prasat pomocí PCR-RFLP (Omelka a kol. , 2001)
Použitá literatura k dispozici u autora.