MOLEKLER BYOLOJ 4 Hafta DNA RNA ve genom

MOLEKÜLER BİYOLOJİ 4. Hafta DNA, RNA ve genom organizasyonu 1

Gen-Genetik Kod-Genetik bilgi Kromozom Nukleus Kromozom Ge n Gen 2

Replikasyon Transkripsiyon DNA r. RNA t. RNA Protein Translasyon Ribozom Santral doğma m. RNA 3

Nükleik asit Kimyası • Nükleotitler: Nükleik asitlerin yapı taşları Fosfa t Pürin ya da pirimidin bazı Pentoz = Nitrojen Baz + Pentoz Şeker + Fosfat grup Pürinler (9 üyeli çift halkalı)– Adenin &Guanin Pirimidinler (6 üyeli tek halkalı–Timin&Sitozin&Urasil 4
 Adenin (A) Pürin Pirimidin Guanin (G) Pürinler Timin")
Nükleik asitlerin bileşenleri (Nitrojen bazları-azotlu bazlar) Adenin (A) Pürin Pirimidin Guanin (G) Pürinler Timin (T) Sitozin (C) Urasil (U) (DNA)Pirimidinler (DNA, RNA) (RNA) DNA RNA 5
 • Ribonükleotitlerde 2’-OH bulunur • Deoxyribonükleotitlerde 2’-H bulunur •")
Nükleik asitlerin bileşenleri (Pentoz şekerler) • Ribonükleotitlerde 2’-OH bulunur • Deoxyribonükleotitlerde 2’-H bulunur • Şekerler azot bazlardan ayırt edilebilmeleri için numaralandırılmış karbon atomları içerirler Bu karbonda oksijen atomu bulunmaz Riboz Deoksiriboz 6

Nükleik asit Kimyası Nükleozitler = Baz + Şeker NMP = nükleozit + 1 PO 4 NDP = nükleozit +2 PO 4 NTP = nükleozit + 3 PO 4 Nükleik asitlerin yapı taşıdır özel NTPs: ATP & GTP Nükleozit Nükleotit Nükleozitlerin isimlendirilmeleri ve genel yapıları 7

Nükleik asit oluşumundaki bağlar • Şekerin C-1’ atomu nitrojen bazla -N glikozidik bağ yapar • Pürin bazı N-9 atomu ile şekerin C 1’ atomu kovalent bağ yapar • Pirimidin bazı N-1 atomu ile şekerin C 1’ atomu kovalent bağ yapar • Nükleotitler şekerin C-2’, C-3’ ve C-5’ atomlarına bağlanabilirler • Ancak, C-5’ konfigürasyonu biyolojik sistemlerdeki en yaygın olan, DNA ve RNA da bulunan mevcut formdur. 8

Nükleozit Difosfatlar ve trifosfatlar, AMP, ADP ve ATP • Ek fosfat grupları nüklozit 5’- monofosfatlara eklenebilir; difosfatlar ve trifosfatlar oluşur • ATP hücre aktiviteleri için en önemli enerji kaynağıdır 5’monofosfat Adenozin 5’-monofosfat (AMP) 5’-difosfat Adenozin 5’-difosfat (ADP) Adenozin 5’-trifosfat (ATP) 9

Polinükleotitler İki mononükleotit arasında bağ yapısında, iki şekere bağlı fosfat grubu yer alır oluşan bağ fosfodiester bağıdır, çünkü fosforik asit her iki taraftaki alkol grubu ( iki şekerdeki OH grubu) ile ester bağı yapar. Aynı bağ, RNA da da bulunur. dinükleotitler & trinükleotitler oligonükleotitler (<20) polinükleotitler (>20) Uzun polinükleotid zincirleri varyasyon sağlamaktadır. � 1000 nt oluşan bir zincir 41000 kombinasyon ile oluşturulabilir. � Levene’nin tetranükleotid hipotezi bu varyasyonu sağlamamaktadır. 10

Şeker-fosfat iskeleti BAZlar 5’ uç Fosfodiester bağları 5' 4' 2' 3' DNA’nın kovalent iskeleti Zincir uzarken yeni eklenen nükleotidin 5. C atomundaki fosfatın OH grubu ile zincirin sonunda yer alan nükleotidin şekerine ait C 3’ atomunda yer alan OH grubu arasında fosfodiester bağı kurulur. 1' Fosfodiester bağları 5' 1' 4' 2' 3' 3’ uç 5' 4' 1' 2' Fosfa 3' t Şeker (deoksiriboz) DNA nükleoti di 11

DNA Modeli: James D. Watson & Francis H. Crick • DNA’nın sarmal çift zincir modelinin sunulması • Bilginin esas iki kaynağı: • 1. Hidrolize olmuş DNA örneğinin baz kompozisyon analizi “Chargaff Kuralı”: #A #T and #G #C. “A strange but possibly meaningless phenomenon”. • 2. X-ray kırınım çalışmaları: Rosalind Franklin & Maurice H. F. Wilkins 12

Watson-Crick Modeli DNA sarmalı Baz, şeker ve fosfat durumu, Hbağları Çap 20 Ao • İki uzun polinükleotit zinciri bir merkez eksen etrafında kıvrılarak, sağ-el ikili sarmal yapısını oluşturur • İki zincir birbirine antiparaleldir; iki zincirin C-5’ ucundan C-3’ ucuna doğru olan yönleri birbirine göre terstir. • Her iki zincirin bazları düzlemsel yapıda olup düzlemlere eksene diktir. Bazlar arasındaki mesafe 3. 4 A olup birbiri üzerine istiflenmiş durumdadırlar ve sarmal içerisinde yer alırlar. • Karşı zincirdeki azotlu bazlar hidrojen bağları ile birbirileri ile eşleşirler. DNA’da sadece A=T ve G C eşleşmesi olabilir (komplementerlik). • Sarmalın her bir tam dönüşü 34 A (3. 4 nm) dir. Her bir zincirde bir dönüşte 10 baz yer alır. • Molekülün herhangi bir bölümünde, eksen boyunca sıra ile daha geniş olan büyük (majör) oluklar ve daha dar olan (küçük) minör) oluklar görülmektedir. • Sarmal 20 A (2 nm) çapındadır. Bir tam dönüş 34 Ao Küçük oluk şeker-fosfat iskeleti Büyük oluk 22 Ao Nitroen Baz çifti Bazların yatay istiflenmesi 3. 4 Ao 13 Merkezi eksen

• Sağ el sarmalının uzaydaki konformasyonu, Watson Crick’in verilerine en uygun olanıdır. DNA sarmalı DNA’daki baz eşleşmesi modelin genetik açıdan en önemli özelliğidir. Bu yerleşim nedeniyle eksen boyunca büyük ve küçük oluklar ortaya çıkar. 14
— yüksek")
DNA’nın formları A-DNA sağ el sarmalı (11 baz/ 1 tam dönüş/çap 23 Å)— yüksek tuz kons. ya da dehidrasyon koşullarında baskındır. A formu su dışında birçok çözeltide oluşan bir formdur. A tipi sarmal RNA nın önemli bir yapısal özelliğidir. RNA çift zincirli bölgeler oluşturduğunda A formunda sarmalı benimser. Bunun nedeni, riboz şekerin 2’ OH grubunun, RNA nın B formunu oluşturmasını engellemesidir. In vitrodan DNA nın A formunu yapmak mümkündür, ancak hücrede bu forma rastlanmamıştır. B-DNA sağ el sarmalı (normal, 10 baz/dönüş, çap 20 Å) (Watson Crick DNA) Fizyolojik koşullarda rasgele bir DNA dizesinde en stabil form B formudur. 15

DNA’nın formları Z formu: Sola doğru dönen heliks yapısındadır. Geçici süre ile oluşan Z formu bazı genlerin ekspresyonlarının regulasyonunda rol alabilir. Bir zincirdeki ardışık GC bazlarında oluşan bir bölgenin komplementeri olan diğer zincirdeki GC bazları ile sola sarımlı bir sarmal oluşturduğu gözlemlenmiştir. DNA zincirleri birbirleri üzerinde saatin aksi yönünde sarılırlar. Z-DNA da her dönüşte 12 baz vardır ve zigzag şeklindedir. Yüksek tuzluluk şartlarında , alkol varlığında oluşturulmuştur, ayrıca ökaryotlarda sitozin metilasyonu devamında normal hücrelerde bu formun oluştuğu tespit edilmiştir. 16
 100 • Isınma ile DNA çift zincirini")
DNA Ergimesi-Denatürasyonu 50 0 Denatürasyon oranı (%) 100 • Isınma ile DNA çift zincirini birarada tutan kovalent olmayan güçler zayıflar ve kırılır • İki zincir birbirinden ayrılır –denaturasyon-ergime • DNA zincirlerinin ½ sinin denatüre olduğu sıcaklık ergime sıcaklığı-melting temperature veya Tm olarak ifade edilir. • DNA nın GC içeriği ergime sıcaklığını önemli oranda etkiler. GC içeriği yüksek DNA nın Tm değeri de yüksektir. • Çift zincirli DNA molekülleri denatüre oldukça OD 260 nm de çok daha fazla ışığı absorbe eder. DNA nın bu özelliğine hiperkromik etki denir. 2 -17

DNA Denatürasyonu • Isının yanısıra, DNA: • Organik çözücüler • Yüksek p. H • Düşük tuz konsantrasyonu ile denatüre olurlar • GC içeriği ayrıca DNA yoğunluğunu etkiler • Doğrudan lineer ilişki • AT baz çiftlerinin GC baz çiftlerinden daha büyük molar hacme sahip olması nedeni ile. 2 -18

DNA Renatürasyonu • İki DNA zinciri ayrıldıktan sonra, uygun şartlarda tekrar bir araya gelebilirler • Bu işlem annealing veya renatürasyon adını alır. • Üç önemli faktör: • Sıcaklık • DNA Konsantrasyonu – Konsantrasyon arttıkça iki zincirin birbirini bulma olasılığı artar • Renatürasyon süresi – süre arttıkça zincir birleşmesi daha fazla olur 2 -19

Hibridizasyon • Birbirine komplementer olan iki tek-zincirli polinükleotit zincirin baz eşleşmesine dayanarak çift-sarmal yapıda nükleik asit molekülünün elde edilmesini sağlayan yapay bir yöntemdir. • DNA: DNA • DNA: RNA • RNA: RNA 20

Farklı büyüklük ve şekilde DNA lar 2 -21

DNA nın katlanması Çift zincirli DNA sarmalının hücre içine paketlenmesi için yüksek bir organizasyona gerek vardır. E. coli kromozomal DNA sı 4600 kbç uzunluğundadır ve 1 mm den uzundur. Bakteri hücrsinden 1000 kat daha uzun olan DNA nın hücre içine sığdırılması için süpersarmal yapı meydana getirilir. Büyük DNA molekülleri hücre içerisinde süpersarmallar oluşturarak bulunurlar Relaks, gevşek süpersarmal 22

DNA nın katlanması • Süperkatlanma-süpersarmal • Negatif katlanma: DNA molekülü kendi ekseni etrafında ters yönünde döndürüldüğünde meydana gelir. • Bu dönüş çift zncirli DNA nın birbiri etrafındaki dönüşünü gvşetir ve zincirlerin çözünmesine yardımcı olur. • Doğada yaygın olarak bulunan DNA molekülleri bu şekilde meydana gelirler. • Pozitif katlanma: DNA çift sarmalının bükülme yönüyle aynı yönde olan katlanma şeklidir. • Bu katlanma çift zincirin daha da sıkı sarılmasına yol açar. Replikasyon sırasında replikasyon çatalının ilerisindeki bölgelerde meydana gelir. 2 -23

DNA nın katlanması • Topoizomeraz II- DNA giraz: • Hem negatif hem de pozitif süpersarmalları DNA yı gevşetmek için görev alan enzimdir. • Reaksiyonun oluşumu ATP bağımlıdır. • Arkelerde ters giraz olup DNA ya ekstra süpersarmalları kazandırmaya yarar. • Bu enzim iki zincirin bir araya geldiği yerde geçici çift zincir kırığı oluşturur • Topoizomeraz I: • ATP gereksinimi olmadan çift zincirli DNA nın tek zincirinde geçici kırık oluşturan enzimlerdir. • DNA yı gevşetmeye yarar. 2 -24

Sıradışı DNA sekonder yapıları • Palindrom – Ters tekrarlar • Ayna tekrarlar • Hairpin (Saç tokası) ve Cruciform (Haç şekli) Yapılar 25

Üçlü sarmal DNA • Çift zincirli DNA ya üçüncü bir DNA zincirinin sarılması ile oluşur. • Daha çok pürin-pirimidince zengin yerlerde meydana gelir ve ayna simetrisi içeren diziler tarafından tercih edilir. • Üçlü sarmal; mevcut DNA içinde tekrarlarda pürin-pirimidince zengin bölgenin katılması ile (intra moleküler tripleks) ya da ayrı bir dizinin katılması ile (inter moleküler tripleks) oluşur. • Üçlü sarmal Watson Crick baz eşleşmesi dışında Hoogsteen AT GC baz eşleşmesi ile olur. 26

Hoogsteen baz eşleşmesi 27

Friedreich ataksisi ve üçlü sarmal DNA • Kas koordinasyonunun kademeli kaybı ve kalp büyümesi ile karakterize edilen nadir bir kalıtsal hastalıkdır. • Hastalık 9. kromozomda ilgili geninn normalde 6 -34 kopyalı olması gereken 5’-GAA-3’ tekrarının 66 -1700 e çıkması ile belli eder. • Bu yaygın GAA motifi üçlü sarmalı tercih eder. Bu üçlü sarmal yapı genin transkripsiyonunu engeller, frataksin proteini azalır, bu mitokondrilerde yüksek demir varlığına yol açar. Aşırı demir yükü oksidatif hasar yol açar. 28

RNA nın çok yönlülüğü • RNA çok amaçlı bir moleküldür. • DNA ya kıyasla daha büyük yapısal değişkenliğe sahiptir. • RNA molekülleri küresel (globular) proteinlere benzer şekilde işlev gören üç boyutlu özgün yapılara katlanırlar. • Katlanma şekilleri diğer moleküller (ligandlar, proteinler, nükleik asitler vb) ile özel etkileşimleri ve onların kimyasal aktiviteleri için temel oluşturur • RNA birçok hücresel süreçte rol almaktadır. 29

RNA çeşitleri • r. RNA: ribozomun temel bileşeni • m. RNA: bir gen ürününü kodlayan genomik DNA dizisinin bir kopyası olup ribozomlara bağlanır • t. RNA: özgün bir amino asit ile «yüklü» küçük bir RNA. t. RNA nın antikodonu ile uygun m. RNA kodonu eşleşerek uygun amino asit ribozoma iletilir. • sn. RNA (RNA işleme): m. RNA yı translasyona hazırlayan pre-m. RNA dan intronların çıkarılması işleminde görev yapar. • sno. RNA: r. RNA nın işlenmesinde rol alır. • mi. RNA: transkripsiyon sonrası gen düzenlenmesinde görev alır. mi. RNA hedef m. RNA dan komplementer bir diziye bağlanır, m. RNA nun yıkımını başlatarak ya da ribozomun translasyon yapmasını engelleyerek geni susturur. • Telomeraz RNA (replikasyon): Doğrusal kromozomlarda replikasyon sırasında uçların kısalmasını önler. 30

RNA nın çok yönlü fonksiyonları • RNA bir «iskele» görevi yapabilir. Yani; sinyal tanıyıcı partiküller (SNP) de olduğu gibi proteinler RNA lar üzerinde düzenli bir biçimde yapılanabililer. • RNA –protein etkileşimleri proteinlerin katalitik aktivitelerini etkileyebilir. Özellikle ökaryotlarda RNA-protein etkileşimleri çok yaygındır. Ribozom da bir ribonükleoproteindir (RNP). • RNP lerde protein enzim olarak iş görür, ancak, substratı hedeflemek ya da bağlanmak için RNA ya ihtiyaç duyar. Ör; telomeraz. • Ribozom ve RNaz da ise RNA katalitiktir, katalizi protein sağlamaz. • RNA katalitik olabilir. Ribozim adlı RNA lar canlı hücrelerde meydana gelen bir çok kimyasal olayı katalizleyebilir. 31

RNA nın çok yönlü fonksiyonları • Kısa RNA lar doğrudan gen ekspresyonunu kontrol edebilirler. RNA interferans, mi. RNA. • RNA kalıtsal materyal olabilir. Birçok virüste genom RNA dır, kendini replike edebilir ya da DNA aracılıı ile replike olabilir. 32

RNA m. RNA: Protein sentezi için gerekli genetik bilgiyi nükleustan sitoplazmaya taşır. t. RNA: m. RNA’daki bilgiyi çözer. r. RNA: Protein sentezi için gerekli hücresel yapı olan ribozomun bileşenidir. (%50 sini oluşturur. ) 33
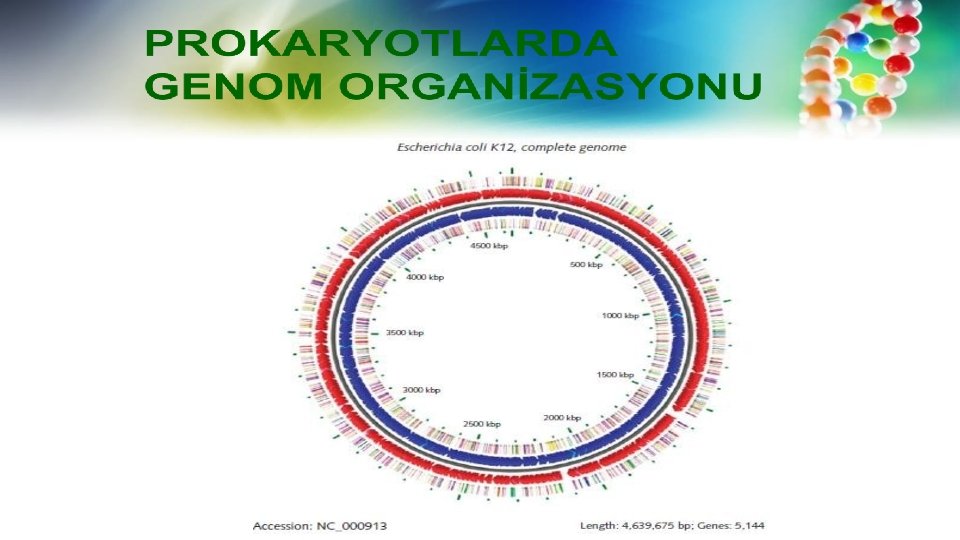

PROKARYOTLARDA GENOM ORGANİZASYONU • Tipik bir prokaryot genomu 0. 6 ila 10 milyon bç uzunluğunda çiftzincirli DNA ya ait tek bir halkasal moleküldür. • Söz gelimi, E. coli K 12 suşunun DNA sı halkasal çift- zincirli olup 4 639 675 bç uzunluktadır. • DNA süperkatlanmış olup histon-benzeri proteinlerle «kromozom» şeklinde paketlenir ve nükleoid olarak adlandırır. • Bazı E. coli hücreleri plazmit içerirler: kısa, genellikle halkasal, 1 kb dan birkaç megabaz uzunluğuna kadar değişen çift zincirli DNA molekülleridir.
 • Süpersarmal (süperkatlanma) • DNA ilmekleri (loops)")
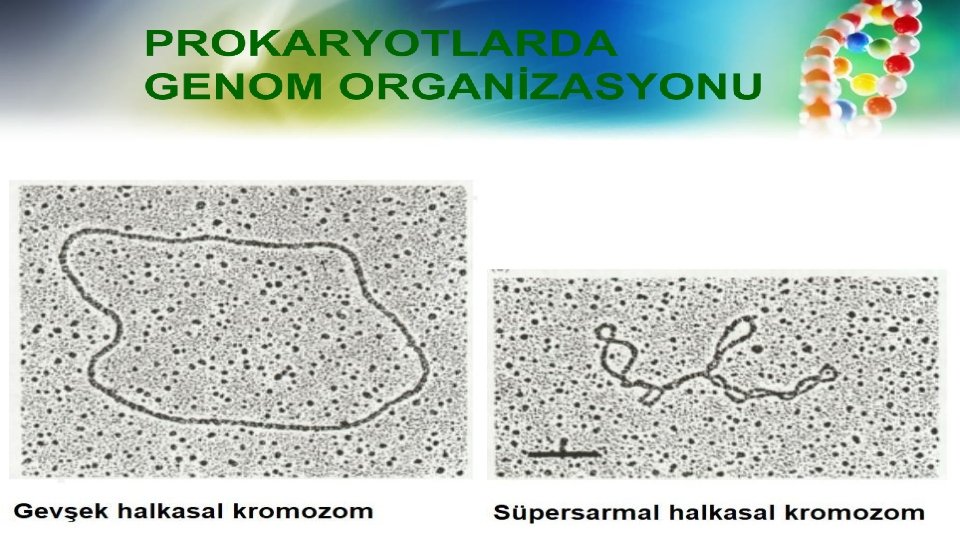
PROKARYOTLARDA GENOM ORGANİZASYONU • Nükleoid (protein-DNA etkileşimi) • Süpersarmal (süperkatlanma) • DNA ilmekleri (loops) Süpersarmal domainlere sahip kromozomal DNA • Genom gevşek ve bir zarla çevrili değildir. • Bakteri genomundaki loop domain paketlenmeye katkıda bulunur. • Genomu bakteriyel bir hücreye sığdırmak için DNA süpersarmal hale gelir.

• Nükleoid 400 bağımsız negatif süpersarmal domainlerine sahiptir. • Süpersarmalların ortalama yoğunluğu 1 dönüş /100 bç’dir. • Her bir domain bir DNA ilmeğinden oluşur, bu ilmeğin uçları, rotasyona izin vermeyen bir şekilde sabitlenmiştir.

• Tek halkasal genom içeren DNA arke ve bakteriler için yaygın olmakla birlikte, bazı prokaryotlarda doğrusal DNA bulunmaktadır. • Ör, Borrelia burgdorferi lyme hastalığının etkeni olup doğrusal DNA ya sahiptir. B. burgdorferi ayrıca çok sayıda plazmit içerir, bunlardan bazıları halkasal, bazıları doğrusaldır. Bazı prokaryotlar birden fazla kromozom içerirler. • Vibrio cholerae, koleraya yol açan patojen bakteri olup 2 961 146 ve 1 072 314 bç uzunluğunda iki halkasal DNA ya sahiptir.
 ye sahiptirler, IS")
• Hepsi olmamakla beraber bazı prokaryot genomları insersiyon sekansları (IS) ye sahiptirler, IS ler hareketli genetik elementler olup ökaryotlardaki transpozonlar ile benzerdir. • E. coli nin 4. 6 Mb lık kromozomu her iki zincirde dağılmış olan yaklaşık 4500 gen içermekte olup intronların ve kısa intergenik bölgelerin olmayışı sebebiyle yüksek kodlama yoğunluğuna sahiptir. • DNA sının %87 si protein kodlarken, %0. 8 i yapısal RNA ları kodlamaktadır; %0. 7 sinin ise fonksiyonu bilinmemektedir.

• Plazmitler kromozomal DNA dan bağımsız olarak kendini eşleyebilirler • Plazmit büyüklüğü 1 - 1000 kbç den daha büyük olabilir.

Kodlama yüzdesi ve ortalama gen yoğunluğu E. coli de gen uzunluğunun dağılımı. Sırasıyla, 7152 ve 8619 bç uzunluklarında olan hipotetik yee. J ve ydb. A, genleri dahil edilmemiştir. Ortalama gen uzunluğu 960 bç dir. Çoğu gen 1500 bç’nden daha kısadır.

• Birçok prokaryotik genom dizilenmiştir. İnsan genomuna göre daha basit içerikte olduklarından tüm yaşam formlarının ortak sorunlarını çözmek için kullanılırlar. Ayrıca, prokaryotik ve ökaryotik metabolizmadaki farklılıklar, söz gelimi, prokaryotlara özgü enzimler enfeksiyona karşı ilaç dizaynında uygun hedeflerdir. Klinik tıptaki önemlerinden bir diğeri ise patojenisite ve antibiyotik direncini nasıl geliştirdiklerini anlamaktır.

• • • Ökaryotik hücrelerdeki genomik bilgi ana nüklear genom ve sitoplazmik organeller olan mitokondri ve kloroplast arasında bölünmüştür. Genom çift zincirli bir DNA dır. Farklı proteinlerle etkileşime girerek kromozom adı verilen çok sayıda doğrusal paketlemeler şeklinde düzenlenmişlerdir.

• DNA histonlarla etkileşir ve nükleozomları oluşturur. • Bir histon oktameri (H 2 A, H 2 B, H 3 ve H 4)X 2 DNA yı sararak nükleozomu oluşturur. • 147 bç DNA her bir histon oktameri etrafına 1. 65 kez sarılır.
 bulunur, ~40 -50")
• • İki nükleozom arasında bağlantı DNA sı (linker DNA) bulunur, ~40 -50 bç. Bu yapı iplik üzerinde boncuklar olarak ifade edilir. H 1 ayrıca, daha ileri yoğunlaşmayı kolaylaştırır. Nükleozomlar solenoid veya 30 nm fiber oluşturmak üzere yoğunlaşırlar. • Nükleozomlar spiral şekilde yoğunlaşırlar.

ÖKARYOTLARDA GENOM ORGANİZASYONU • Kromatinde Ökromatin ve Heterokromatin bölgeler bulunur

ÖKARYOTLARDA GENOM ORGANİZASYONU • Ökromatin: Hücre döngüsü boyunca yoğunlaşan ve açılan bölgeler olup normalde aktif bölgeleri (gevşek paketlenme) temsil ederler, Çünkü: Transkribe olan bölgelerdir ve çok fazla tekrarlı diziler içermezler • Heterokromatin: Hücre döngüsü süresinde yoğun (kapalı) olan bölgelerdir. Bu bölgeler gen olmayan ya da transkripsiyona uğrayan genlerin olmadığı yerlerdir.

İnsan kromozmunda DNA içeriği

ÖKARYOTLARDA GENOM ORGANİZASYONU Metafaz kromozomları: • Sentromer ile ayrılmış iki kol (kısa olan p, uzun olan q) • Sentromer • Telomer den oluşur.

• Electron micrograph of a proteindepleted human metaphase chromosome, showing the residual chromosome scaffold and loops of DNA. Individual DNA fibers can be best seen at the edge of the DNA loops. Bar 2. (From Paulson JR, Laemmli UK [1977] The structure of histone-depleted metaphase chromosomes. Cell 12: 817– 828. Reprinted by permission of the authors and Cell Press. )

• Kromozomların en yoğun formu metafaz kromozomlarıdır. • Metafaz kromozomları tarafından temsil edilen tüm genom karyotip adını alır. • Belirli bir ökaryotik hücrede kromozom setleri sayısı ploidi seviyesini vermektedir.

• Genomun kromozomlara paketlenmesi
: Bir set kromozom içeren hücrelerdir. Örneğin, insan")
ÖKARYOTLARDA GENOM ORGANİZASYONU • Haploid (1 N): Bir set kromozom içeren hücrelerdir. Örneğin, insan gamet hücreleri • Diploid (2 N): Bir kromozoma ait iki set kromozom içeren hücrelerdir. Ör, insan somatik hücreleri.

Haploid bir insan hücresi 23 nüklear kromozomdan, bir mitokondri kromozomundan meydana gelirken 3. 2 milyardan fazla DNA baz çiftine sahiptir. • Karyotik genom doğrusaldır ve Watson Crick ikili sarmal yapısal modeline uygun yapıdadır. DNA üç seviyeli paketlenme (ipe dizili boncuklar, nukleozomlar, ilmek&domainler)ile kromozomları oluştururlar. • Ökaryotik genomlar, kodlama dizisini ve RNA parçasının işlevselliğini temsil eden araya giren dizileri belirtmekte olan protein kodlayan genlere ait eşsiz özellikte ekzon- intron organizasyonuna sahiptirler. •

• Ökaryotik kromozomlar tekrarlı DNA ların varlığı ile kompleks bir organizasyona sahiptirler. • Tekrarlı DNA dizileri pek çok kez ökaryotik kromozomda tekrarlanırlar.

Ökaryotik genom konfigürasyonu 57


• Protein kodlayan diziler önce m. RNA ya transkribe olan ardından polipeptitlere translasyon gerçekleştirilen DNA dizileridir. • Genomun tam protein kodlayan gen kapasitesi, ekzomlar içinde bulunur (RNA splaysing ile intronların çıkarılması sonrası transkribe edilen ekzonlardan oluşan genom kısmı) ve ekzonlar tarafından kodlana DNA dizileri proteinlere transle edilir. • Açık Okuma Çerçeve (ORF) lerden oluşur. Bu diziler proteinler/peptitleri kodlama potansiyeline sahiptir. t consists of ORF (Open reading frame). These are the reading frame that has the potential to code for the proteins/peptide. UAA, UAG, ve UGA dur kodonları bulunmayan bir dizidir. AUG translasyonun nnerede başlayacağını belirtebilir.

• Promotorlar genellikle bir genin transkripsiyon başlangıç noktasının 200 bç yukarısında konumlanan kısa dizilerdir. Farklı bileşenlere ayrılabilirler. – Proksimal promotör bölgesi, çekirdek (öz) promotorun hemen yukarısında, genellikle 50 ila -200 bç arasında bulunan sekanstır. – ENHANCERlar: Memeliler gibi kompleks ökaryotlarda özellikle yaygın olan, ancak maya gibi basit ökaryotlarda bulunmayan veya çok zayıf bir şekilde temsil edilen pozitif transkripsiyonel kontrol elemanlarıdır. – SILENCERlar: transkripsiyon seviyesini azaltmaya yararlar. – SINIR ELEMENTLERİ (INSULATORS): Bu DNA bölgeleri, genellikle 0. 5 kb ila 3 kb arasında değişen, enhancer veya silencer ların etkilrinin yayılmasını engelleyici işlevlere sahiptirler. – RESPONSE ELEMENTS YANIT ELEMANLARI spesifik eksternal uyaranlara yanıt olarak transkripsiyonu modüle eder. Genellikle promotör elemanların yukarısında kısa bir mesafe bulunurlar (genellikle transkripsiyon başlangıç bölgesinin 1 kb'si içinde). Bu gibi çeşitli elementler spesifik hormonlara veya siklik AMP gibi hücre içi ikinci habercilere yanıt verir.
 proteine çevrilmeyen RNA molekülüdür. Daha az sıklıkla protein")
• Kodlamayan RNA (nc. RNA) proteine çevrilmeyen RNA molekülüdür. Daha az sıklıkla protein kodlamayan RNA (npc. RNA), mesajcı olmayan RNA (nm. RNA) ve fonksiyonel RNA (f. RNA) olarak adlandırılır. Fonksiyonel bir kodlamayan RNAyı transkribe eden DNA dizisi çoğunlukla RNA geni olarak ifade edilir.
 22 nt")
RNA sınıfı Fonksiyon r. RNA 150 -5000 nükleotid micro. RNA (mi. RNA) 22 nt Gen regülasyonunda özellikle ost transkripsiyonel regülasyonda önemli görevleri vardır t. RNA 70 -80 nükleotid Piwi-bindin RNA (pi. RNA) 24 -31 nt Tekrarlarda kökenlenir, sadece germ hattu hücrelerinde ifade edilie, aşırı transpozon aktiitesini sınırlar. Küçük çekirdek RNA (sn. RNA) 60 -360 nt Splaysozom yapısında bulunur Endojen kısa arayagiren RNA (endosi. RNA) 21 -22 nt Psödogenler, ters tekrarlarda kökenlenr, somatik hücrelerde gen regülasyonunda görev alır. Küçük çekirdekçik RNA (sno. RNA) 60 -300 nt r. RNA nın olgunlaşması Uzun kodlamayan regülatör RNA (long RNA) >1 kb Gen ifadesinin düzenlenmesinde rol oynar, bazıları Xinaktivasyonuna katılır, bazıları antisens regülatörlar olarak işlev görür Küçük Kajal badi RNA (sca. RNA) Belirli sn. RNA ların çekirdekte kaja badide olgunlaşması Küçük sitoplazmik RNA lar 80 -500 nt RNA ribonükleazlar 260 -320 nt Nükleus ve mitokondride pre t. RNA yı keser

• İntron terimi bir gendeki DNA dizisi hem de RNA transkriptinde olarak ifade edilir En az dört ayrı intron sınıfı belirlenmiştir. • Translasyon olmayan bölge (UTR) bir m. RNA zincirindeki kodlama dizisinin bölgeleri ifade eder. 5' tarafında bulunuyor ise 5' UTR (veya lider dizi), 3' tarafta ise 3' UTR (ya da fragman dizisi) olarak adlandırılır. karşılık gelen dizi

Gen fragmentleri sadece ekzonları içeren gen parçalarıdır. c. DNA dan oluşurlar.

• Hücrede gen ifadelerini ya da protein kodlama yeteneklerini kaybetmiş, genlerin işlvsiz akrabalarıdır. Psödogenler çoğunlukla, ürünü organizmanın yaşaması için gerekli olmayan bir gende çok sayıda mutasyonnun birikmesi sonucunda oluşurlar. DNA dizisine göre, iki tipi vardır: • İşlenmiş psödojen: Protein kodlayan bir genin tüm normal kısımlarına sahiptirler, ancak başlangıçta varsayılan DNA kodu hatalarına dayanarak etkisiz oldukları düşünülmüştür. • İşlenmemiş psödojen: İntronları olmayan psödogenlerdir.
; uzunluğu 200 nükleotidi aşan bir tür kodlamayan")
• Uzun kodlamayan RNA (lnc. RNA); uzunluğu 200 nükleotidi aşan bir tür kodlamayan RNA (nc. RNA)’dır. lnc. RNAlar memeli transkriptomunda nispeten boldur ve kromatin-odifiye edici enzimlerin katılımı yoluyla gen transkripsiyonunun regülasyonunu kapsayan, çeşitli hücresel fonksiyon ile rol oynamaktadır.


• İntergenik bölge genler arasında konumlanan DNA dır. İntergenik bölgeler kodlamayan DNA birimleridir. Bazı intergenik bölgeler yakınında oldukları genleri kontrol ekmek üzere görev üstlenirken, birçoğu şuan bilinen fonksiyona sahip değillerdir. Bazen junk DNA olarak isimlendirilirler. • İnsanlarda, intergenik bölgeler genomun %75 ini oluştururken bu sayı bakterilerde (%15) ve mayalarda (%30) daha azdır. • İntergenik bölgeler intragenik bölgelerden (intronlar) farklıdırlar. İntronlar kısa gen içerisinde bulunan kodlamayan bölgelerdir, özellikle ökaryotik genlerde bulunurlar. • Promotor ya da enhancer gibi işlevsel açıdan önemli elementleri içermezler. Ayrıca, intergenik bölgeler kodlamayan RNA lar gibi henüz tanımlanmamış genler içerebilir. Regülatör işlevleri olduğu düşünülmektedir.

B 2. ORTA/YÜKSEK TEKRARLI • Yüksek oranda tekrarlanan kodlamayan insan DNA sı çoğunlukla basit (1 -10 nükleotid) veya orta derecede veya yüksek sayıda (10 ila yüzlerce nükleotid) olabilen ardışık tekrarlı sekans bloklarından meydana gelir. Bireysel bloklar birkaç ya da çok fazla sayıda farklı kromozoma bölgesinde (lokasyonunda) meydana gelebilir. • Satellit DNA: Satellit (uydu) minisatellit DNA nın büyük çoğunluğunda olduğu gibi transkripsiyonel olarak inaktiftir, mikrosatellit DNA larda ise önemli bir yüzde kodlayan DNA da lokalize olmuştur. – Bir veya daha fazla nükleotit paterni tekrarlandığında ve tekrarlar doğrudan birbirine bitişik olduğunda DNA'da tandem tekrarları meydana gelir.

• Minisatellit DNA: Nükleer genomun önemli bölümleri üzerine dağılmış olan minisatellit DNA ardışık olarak tekrarlanan DNA dizisinin orta büyüklükte dizilim (blok) koleksiyonundan oluşur. – 10 -60 bç uzunluğundaki belirli DNA motiflerinin tipik olarak 5 -50 kez tekrarlandığı repetitif DNA sistemidir. Minisatellitler insan genomunda 1000'den fazla yerde bulunur ve yüksek mutasyon oranları ve popülasyondaki yüksek çeşitliliği nedeniyle dikkat çekici bölgelerdir. – Minisatelliler kromozomların telomer ve sentromerlerinde belirgindirler.

• Mikrosatellit DNA: Mikrosatellit DNA, aynı zamanda basit dizi tekrarları (simple sequence repeats (SSR)) olarak adlandırılmaktadır. Mikrosatellit DNA genellikle 10 bç’nden daha kısa ardışık tekrarlı dizilere ait ardışık küçük bloklardır (array). 60 Mbazdan daha fazlasına karşılık gelerek genom boyunca dağılmışlardır. Bu büyüklük genomun %2 sine karşılık gelmektedir. Ve çoğu kez replikasyon kayması ile ortaya çıktıkları düşünülmektedir.
 gelir, fakat ayrıca bazı protein kodlayan")
• Serpiştirilmiş tekrarlar başlıca transposable elementlerden (TE) gelir, fakat ayrıca bazı protein kodlayan gen alellerini ve psödogenleri içerirler. Transpoze edilebilir elemanlar, hücre içindeki başka bir bölgedeki genom ile bütünleşebilir. TE'lerin yüksek ökaryotların genom evrimi üzerinde önemli bir itici güç olduğuna inanılmaktadır. TE'ler iki kategoriye ayrılabilir: Sınıf 1 (retrotransposzonlar) ve Sınıf 2 (DNA transpozonları).
 çok başarılı transpozonlardır.")
• LINES: • LINEs (long interspersed nuclear elements-uzun serpiştirilmiş-dağınıknüklear elementler) çok başarılı transpozonlardır. Fareler de dahil olmak üzere diğer memelilerde ortaya çıkan nispeten uzun bir evrim geçmişine sahiptirler. • Otonom transpozonlar olarak, temel transkriptaz dahil olmak üzere retro transpozisyon için gerekli tüm ürünleri yapabilirler.
 100")
• SINES: • SINEs (kısa serpiştirilmiş nüklear elementler; short interspersed nuclear elements) 100 -4000 bç uzunluğundaki retrotranspozonlardır. • Memeli genomunda bulunan bu elementler çok fazla kopyalar halinde bulunarak çeşitli sepiştirilmiş DNA ailelerini oluştururlar. LINE lardan farklı olarak SINE ler herhangi bir protein kodlamazlar ve bağımsız olarak hareket edemezler. Bununla birlikte, SINE ve LINElar 3’ uçlarındaki dizileri paylaşırlar, ve SINE lerin komşu LINE larla hareket ettikleri gösterilmiştir. • Insan Alu ailesi kopya sayısı bakımından en belirgin SINE ailesidir. İnsan genomunda en bol bulunan sekanstır. Ortalama her 3 kb da görülürler.

• LTR ELEMENTLERİ: • LTR transpozonları otonom ve otonom olmayan retrovirüs-benzeri elementleri içermekte olup gerekli transkripsiyonel elementleri kapsayan uzun terminal tekrarlarla (LTRs) çevrilidirler.

• DNA TRANSPOZONLARI: • Sınıf II Transpozon elementlerinin kes-yapıştır transpozisyon mekanizması bir RNA araürününü içermez. Transpozisyonlar çeşitli transpozaz enzimleri tarafından katalizlenir. Bazı transpozazlar DNA da herhangi bir hedef bölgeye spesifik olmadan bağlanırlarken, bazıları ise spesifik DNA dizi hedef lerine bağlanırlar. • DNA transpozonları terminal ters tekrarlara sahiptirler ve • transpozisyonu düzenleyen bir transpozaz kodlarlar. İnsan genomunun yaklaşık %3 üne karşılık gelirler. Ritüel olarak yerleşik insan DNA transpozon sekansları artık aktif değildir; bu yüzden transpozon fosilleridir.

KAYNAKLAR • Modern Moleküler Biyoloji Prof. Dr. Nihat DİLSİZ, Palme Yayıncılık, 2017 • Moleküler Hücre Biyolojisi, Prof. Dr. Hasan Veysi GÜNEŞ , 2014, istanbul Tıp Kitabevi • Moleküler Hücre Biyolojisi, 6. baskıdan çeviri, 2011, Palme Yayıncılık, Çeviri editörleri: Prof. Dr. Hikmet Geçkil, Prof. Dr. Murat Özmen, Prof. Dr. Özfer Yeşilada • Temel Moleküler Biyoloji, 2. baskıdan çeviri, Çeviri Editörü: Prof. Dr. Ali Osman Beldüz, 2014 77
- Slides: 77