MOLEKLER BYOLOJ 1 Gen ifadesinin dzenlenmesi Prokaryotik sistemlerde

MOLEKÜLER BİYOLOJİ 1

Gen ifadesinin düzenlenmesi

Prokaryotik sistemlerde gen ifadesinin düzenlenmesi

Genel Özellikler Prokaryotik genler polisistronik sistemlerdir, dolayısıyla birbiriyle ilişkili birçok gen tek bir transkript oluşturmak üzere bir araya gelebilir ve operon adını alırlar. Gen ifadesinin düzenlenmesi genellikle negatif şekildedir. Uyarıcılar baskılamayı kaldırmak için kullanılır. Negatif kontrol: Bir çeşit regülatör molekül (represör) tarafından engellenmediği sürece genetik ifade gerçekleşebilir. Pozitif kontrol: Sadece regülatör molekül (aktivatör), RNA sentezini doğrudan uyarırsa transkripsiyon gerçekleşir.

Transkripsiyonun düzenlenmesi Operon transkripsiyonun düzenlenmesi için koordinasyon birimidir. Transkripsiyonun başlaması düzenleme için en önemli noktadır. Ayrıca translasyon da düzenlenebilir.

LAKTOZ METABOLİZMASI lac operonun yapısı Regülatör bölge Yapısal Operatör Promotor CAP-bağlanma bölgesi Regülatör gen bölge Transasetilaz Permeaz -galaktozidaz

Laktoz operonunun NEGATİF düzenlenmesi lac I geninin kodladığı protein Lac operonunun promotorundaki operatör bölgeye bağlanır ve RNA polimerazın ilerlemesini engeller. Lac l, Lac represör (baskılayıcı) protein olarak da bilinir. Represör protein, allosterik (şekil değiştirme) olarak kontrol edilebilen bir proteindir ve “inducer” (laktoz) olarak bilinen bir molekül için bağlanma bölgesine sahiptir. Bağlanma olduğunda, baskılayıcının bağlanma özellikleri değişir bu da operatorün açık kalmasına neden olur
 NEGATİF İNDÜKSİYON The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland")
Laktoz (-) NEGATİF İNDÜKSİYON The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland (MA): Sinauer Associates, Inc. ; c 2000
 NEGATİF İNDÜKSİYON The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland")
Laktoz (+) NEGATİF İNDÜKSİYON The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland (MA): Sinauer Associates, Inc. ; c 2000

Laktoz operonunun POZİTİF düzenlenmesi

Pozitif düzenlenme. Uyarılabilen gen ifadesi • Ortamda glukoz olduğu zaman bakteri bu üç enzimi üretmez. Bakteri bu enzimleri ortamda glukoz olmayıp laktoz bulunduğu durumlarda üretir.
 zayıf bir promotöre sahiptir, bazal bir ifade düzeyine")
lac operon • lac operon (TTTACA/TATGTT) zayıf bir promotöre sahiptir, bazal bir ifade düzeyine sahiptir. • CAP (Catabolite gene activator protein) bağlanma bölgesi -60 bölgesindedir. • CAP hem DNA’ya hemde c. AMP’ye bağlanabilecek uygun nükleotid dizilerine sahiptir.
![• Glukoz c. AMP oluşumunu engeller. • Glukoz ortamda olduğu zaman, [c. AMP]](http://slidetodoc.com/presentation_image_h2/158e040b6cc51471f460d07fb68c61b2/image-13.jpg "• Glukoz c. AMP oluşumunu engeller. • Glukoz ortamda olduğu zaman, [c. AMP]")
• Glukoz c. AMP oluşumunu engeller. • Glukoz ortamda olduğu zaman, [c. AMP] konsatrasyonu düşüktür. Glukoz tamamen tüketilince, [c. AMP] konsantrasyonu artmaya başlar. The CAP-c. AMP birliği oluşur, ve bu birlik lac operonun CAP binding site (Cap bağlanma bölgesine) bağlanır.

Durum 1 Glukoz yok Laktoz yok, lac geni ifade edilmez.

• lac. I geni kendi promotörüne sahiptir ve bunun aktivitesi sonucunda Lac. I repressör üretilir. • Tetramerik Lac repressör lac operator bölgesine Olac bağlanır. • Bu bağlanma RNA polimerazın DNA üzerindeki hareketini engeller böylece lac. Z, lac. Y, and lac. A genleri ifade edilmez.

Durum 2 Glukoz yok Laktoz ortamda bulunduğu zaman, lac. Z, lac. Y, ve lac. A genleri ifade edilmeye başlanır. galactosidase

• Galaktosidaz bazal durumda çok az ifade edilir. • Ortamda laktoz olduğu durumlarda hemen allalaktoz ve galaktoza dönüştürülür ve repressöre bağlanır. • Repressör artık operatör bölgesine bağlanamaz ve lac genleri ifade edilebilir. • Galaktoz, Allalaktoz ve İPTG uyarıcı (inducer) olarak adlandırılır.

uyarıcılar
![Durum 3 Glukozun bulunduğu durumda, [c. AMP] konsantrasyonu düşüktür ve CAP-c. AMP oluşamaz, Laktoz](http://slidetodoc.com/presentation_image_h2/158e040b6cc51471f460d07fb68c61b2/image-19.jpg "Durum 3 Glukozun bulunduğu durumda, [c. AMP] konsantrasyonu düşüktür ve CAP-c. AMP oluşamaz, Laktoz")
Durum 3 Glukozun bulunduğu durumda, [c. AMP] konsantrasyonu düşüktür ve CAP-c. AMP oluşamaz, Laktoz var; lac operonun ifade düzeyi oldukça düşük seviyelerde gerçekleşir. Bazal ifade

Durum 4 Glukoz yok Laktoz var; CAP-c. AMP birlikteliği CAP bölgesine bağlanarak lac genlerinin aktivasyonunu sağlar.

Koordineli ifade şekli Laktoz Glukoz - Glukoz + Laktoz +
 OPERONU trp operonu bazal düzeyde konstitif olarak (sürekli) ifade edilen")
BASKILANABİLİR SİSTEM: TRİPTOFAN (Trp) OPERONU trp operonu bazal düzeyde konstitif olarak (sürekli) ifade edilen genlerden biridir. trp operonun yapısındaki genler triptofan amino asidinin sentezinde görev alan 5 farklı enzimi kodlarlar. www. nature. com/. . . /images/nbt 1204 -1502 -F 1. jpg

Trp Operonunun Düzenlenmesi 1. Represör/operatör ilişkisi ile 2. Sentezine başlanmış olan transkriptin terminasyonu ile (attenüasyon)

• trp repressör geni ifade edilebilir ama operatör bölgeye bağlanmaz. • TRP gereğinden fazla üretildiği zaman repressör Trp ile birliktelik oluşturarak operatör bölgesine bağlanır ve Trp sentezini durdurur.
 • Hücrede trp varsa, trp’nın bir kısmı")
Trp Operonunun Düzenlenmesi Represör/operatör ilişkisi: Trp (+) • Hücrede trp varsa, trp’nın bir kısmı serbest olan trp. R geni ürünü REGÜLATOR PROTEİN’e (R) bağlanır ve onu aktive eder. • R-trp kompleksi trp operatörüne karşı yüksek affiniteye sahiptir. İlişki kurduklarında, RNA polimerazın ilerlemesini bloke ederler ve transkripsiyonu durdururlar. Operon “kapatılır”.
 Hücrede Trp konsantrasyonu çok azaldığında, Trp,")
Trp Operonunun Düzenlenmesi Represör/operatör ilişkisi : Trp (-) Hücrede Trp konsantrasyonu çok azaldığında, Trp, regulatör proteinden ayrılır, R proteini operatöre karşı olan affinitesini kaybeder ve ayrılır. RNA polimeraz ilerleyerek, Trp sentezinden sorumlu 5 genin transkripsiyonunu yapar.
 düzenlenme")
Trp operonunun düzenlenmesi: Sentezine başlanmış olan transkriptin terminasyonu ile (attenüasyon) düzenlenme
 düzenlenme Ortamda trp miktarı")
Trp operonunun düzenlenmesi: Sentezine başlanmış olan transkriptin terminasyonu ile (attenüasyon) düzenlenme Ortamda trp miktarı fazlaysa ve hızlı translasyona yetecek t. RNAtrp varsa, ribozom bölge 1 -2’yi kendiliğinden örter. 3 -4 bölgeleri ise İLMEK yapar. Sentezi başlamış olan transkriptin RNA polimeraz ile ilişkisi kesilerek transkripsiyon durdurulur. YÜKSEK Triptofan Miktarı The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland (MA): Sinauer Associates, Inc. ; c 2000
 düzenlenme Ortamda hazır bulunan")
Trp operonunun düzenlenmesi: Sentezine başlanmış olan transkriptin terminasyonu ile (attenüasyon) düzenlenme Ortamda hazır bulunan triptofan yetersizse, uygun t. RNAtrp oluşturulması gerekir. Bu sırada ribozom, t. RNAtrp oluşturulmasını beklerken bir gecikme olur ve bu arada 2 -3 bölgeler İLMEK yapar. 3. Bölge, 4 ile İLMEK yapamaz ve RNA polimeraz transkripsiyona devam eder. DÜŞÜK Triptofan Miktarı The Cell - A Molecular Approach Cooper, Geoffrey M. Sunderland (MA): Sinauer Associates, Inc. ; c 2000

Trp operon

Yavaşlatma mekanizması • trp operonu yavaşlatma mekanizması kullanılarak translasyon seviyesinde kontrol edilebilirler.

Ökaryotlarda gen ifadesinin düzenlenmesi

Yapısal özellikler Büyük genom: 3 x 109 bps, 20000 -25 000 gen Monosistron yapı Tekrarlanmış diziler: farklı büyüklük ve farklı sıklıkta. Çoğunlukla ters tekrarlardan oluşmakta. Mozaik gen yapısı: intron ve exonlar

Düzenlemenin özellikleri 1. RNA-pol: Farklı RNA’lar için 3 farklı RNA polimeraz (I, II, and III) 2. Farklı kromozom yapıları • Çok hassas bölge yapısı • Baz modifikasyonu • İzomer dönüşümü • Histon değişiklikleri

3. Pozitif düzenleme 4. Transkripsiyon ve translasyon ayrılmış durumdadır 5. Transkripsiyon sonrası modifikasyonlar oldukça fazladır 6. Hücre içi ve hücreler arası sinyaller yardımıyla gen ifadesinin düzenlemesi görülür.

Gen ifadesinin düzenleyici unsurlar 1. DNA nın paketlenmesi/açılması 2. Transkripsiyon-Transkripsiyon başlangıcı gen ifadesinin kontrol edilmesindeki ana noktadır. 3. RNA transportu (taşınımı) 4. m. RNA yıkımı 5. Post-transkripsiyonel modifikasyonlar 6. Translasyon 7. Post-translasyonel modifikasyonlar
")
Gen İfadesinin Düzenlenmesi - ppt indir (slideplayer. biz. tr)

1. Kromatin düzenlenmesi Kromatin yapısı transkripsiyonun kontrolünde önemli bir aşamadır. Kromatinin büyük kısmı bazı hücrelerde transkripsiyonel olarak inaktif durumda iken diğerlerinde aktif durumdadırlar. Ör; beta globin gen kümesi retikülositlerde «aktif» kromatinde, kas hücrelerinde «inaktif» kromatindedir.

Nükleozomlar-zincir üzerinde boncuklar

Paketlenme ve gen ifadesinin kontrolü Heterokromatin bölge: Koyu DNA bölgeleri sıkı paketlenme; genler kapalı-transkripsiyon yok. Ökromatin bölge: Daha açık DNA bölgeleri, gevşek paketlenme; genler açık-transkripsiyon var. Paketlenme seviyesi gen transkripsiyonunu kontrol eder.

DNA Metilasyonu transkripsiyon faktörlerini bloke eder. Prokaryot ve ökaryotlarda gen ifadesinin baskılanması için genel bir mekanizmadır. DNA metiltransferaz enzimi; S-adenozil-metiyonin’den bir metil grubunun Sitozin’in 5. karbon atomuna transferini katalizler. Sürekli bir gen baskılama modifikasyonu olarka ifade edilebilir. Ör; inaktif memeli X kromozomu olan bar cisimciği. www. cellscience. com/reviews 7/Taylor 1. jpg

Histon asetilasyonu Histonların asetilasyonu DNA yı açar. Histonlara gevşek sarılma genleri açar, transkripsiyon gerçekleşir. Histon proteinlerindeki konformasyonel değişim transkripsiyon faktörlerinin DNA ya bağlanmalarını sağlar. chemistry. gsu. edu/images/nucleosome. jpg

2. Transkripsiyonun başlaması Cis acting elementler DNA molekülüne spesifiklerdir ve bir ya da birden fazla genin transkripsiyonunu düzenlerler. Genellikle korunmuş DNA bölgelerine sahiptirler. Promotör: DNA dizisinde RNA polimerazın ve transkripsiyon faktörlerinin bağlandığı yakın kontrol dizileridir: TATA kutusu, CAAT kutusu, ve GC kutusu

Promotor: TATA kutusu • Dizi: TATAAAA • Lokasyon: - 25 ~ - 30 bp • Fonksiyonu: RNA polimerazın bağlanması içine gerekli olan TFII D için bağlanma bölgesi görevi görür. Transkripsiyonel başlangıcın sıklığını ve doğruluğunu kontrol eder.

Promotor: CAAT kutusu Dizi: GCCAAT Lokasyon: ~ -70 bp Fonksiyon: CTF 1 (CAAT-binding transcription factor) ve C/EBP (enhancer binding protein) için bağlanma bölgesi görevi görür.

Promotor: GC kutusu • Dizi: GGGCGG • Lokasyon: -30 ~ -110 bp • Fonksiyon: Sp 1 proteini için bağlanma bölgesidir.

Enhancer-Arttırıcı Transkripsiyonun zamana ve yere bağlı olarak spesifikliğini ayarlayan ve promotör aktivitesini arttıran DNA dizisidir. «artan transkripsiyon» Aktivatör proteinlerin DNA ya bağlandıkları uzak kontrol dizileridir.

Susturucu-silencer Negatif düzenleme elementidir. Spesifik proteinle etkileştiği zaman transkripsiyonu durdurur.

Enhancer etki mekanizması Uzak DNA dizileri olan enhancerlara aktivatör proteinler bağlanarak tarnskripsiyonu tetiklerler. Silencer proteinler ise enhancer dizilerine bağlanırlar ve gen transkripsiyonunu bloke ederler.
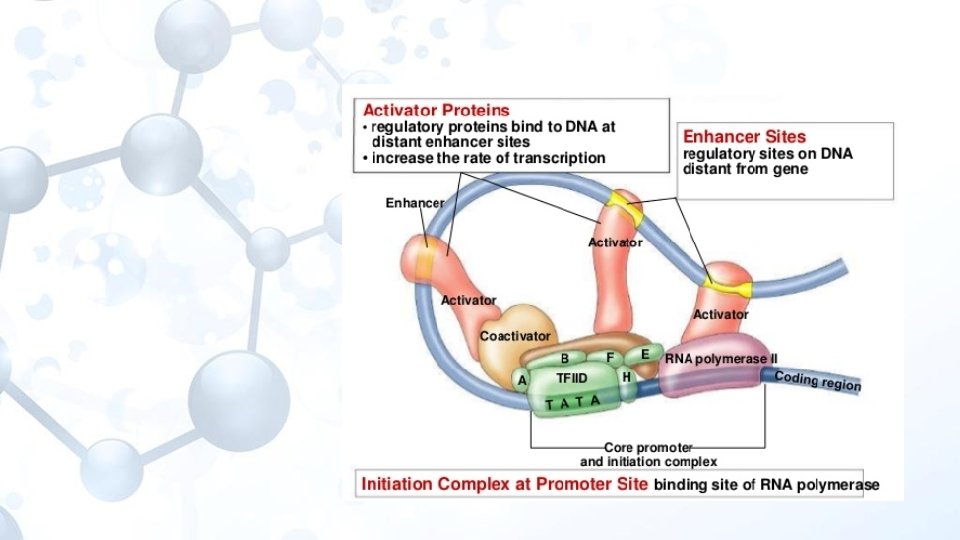

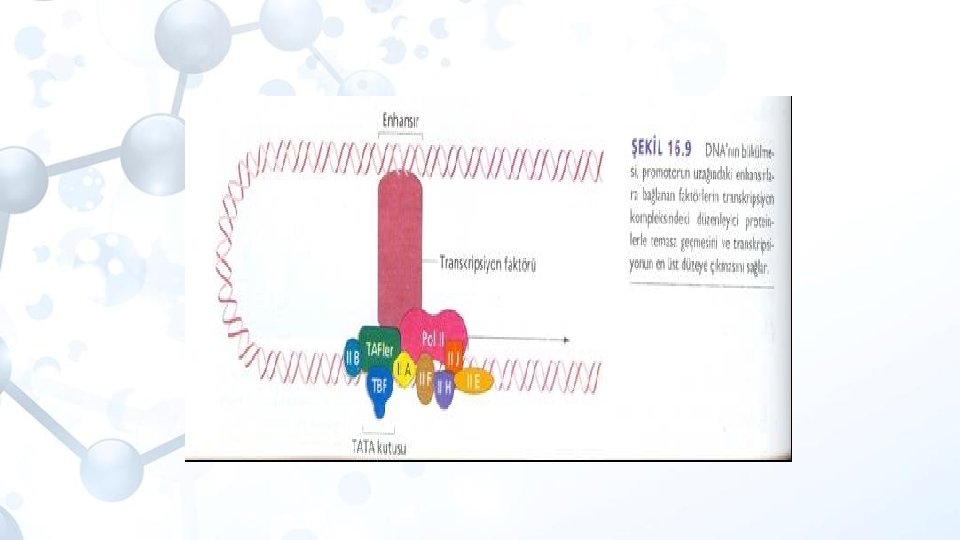
Enhansırlar ve promotorların karşılaştırırsak; Promotorlar transkripsiyon için temel düzeyde gereklidirler. Enhansırlar daha üst düzey kontrol aşamasında etkindirler. Enhansırlar doku ve hücreye özgül iken promotorlar daha geneldir. RNA polimerz promotora, transkripsiyon faktörleri ise enhansırlara bağlanırlar. Enhancera TF bağlanması kromatin konfigürasyonunu değiştirirek RNA polimerazın promotorla birleşmesine yardımcı olur.

Trans acting elementler: Transkripsiyon faktörleri Dolaylı olarak cis-acting elementlere bağlanan ve transkripsiyonun başlamasını düzenleyen proteinlerdir. Genel transkripsiyon faktörleri: transkripsiyonda görev alırlar Özel transkripsiyon faktörleri: Transkripsiyon aktivatörleri-pozitif faktörler EBP (enhancer bağlanma proteini) Transkripsiyon engelleyiciler-represörler
’lar.")
Pozitif faktörler de ikiye ayrılırlar; • Gerçek aktivatör transkripsiyon faktörleri • Antirepresör (karşı baskılayıcı)’lar. Bu proteinler, genlerin ne zaman ve nerede ifade olacağını ve transkripsiyonun oranını kontrol ederler. Aktivatörler, kuvvetlendirici bölgeye (enhensır) bağlanır ve promotordaki transkripsiyon kompleksi ile etkileşerek transkripsiyonun başlama oranını yüz kat artırabilirler. Gerçek aktivatörler promotorda oluşan transkripsiyon faktörleri ile bağlantılı iş yapar. Antirepresörler ise kromatin yapısını değiştirerek diğer faktörlerin bağlanmasını sağlarlar.
 denilen kompleksleri oluşturan modüler proteinlerdir. Enhensozom oluştuktan")
Aktivatörler enhensır DNA dizilerine bağlanarak enhensozom (enhanceosome) denilen kompleksleri oluşturan modüler proteinlerdir. Enhensozom oluştuktan sonra, buradaki proteinler, transkripsiyon kompleksindeki proteinlerle etkileşirler. Aktivatörlerde bu etkileşimi yapmak için iki işlevsel bölge (domain: proteinlerde özgül işlevlerin yapıldığı amino asit kümesi) mevcuttur.

TF’lerinin genel yapısı DNA-bağlanma bölgesi Aktivasyon bölgesi Protein-protein etkileşim bölgesi
 bulunur. Sarmal-dönüş-sarmal")
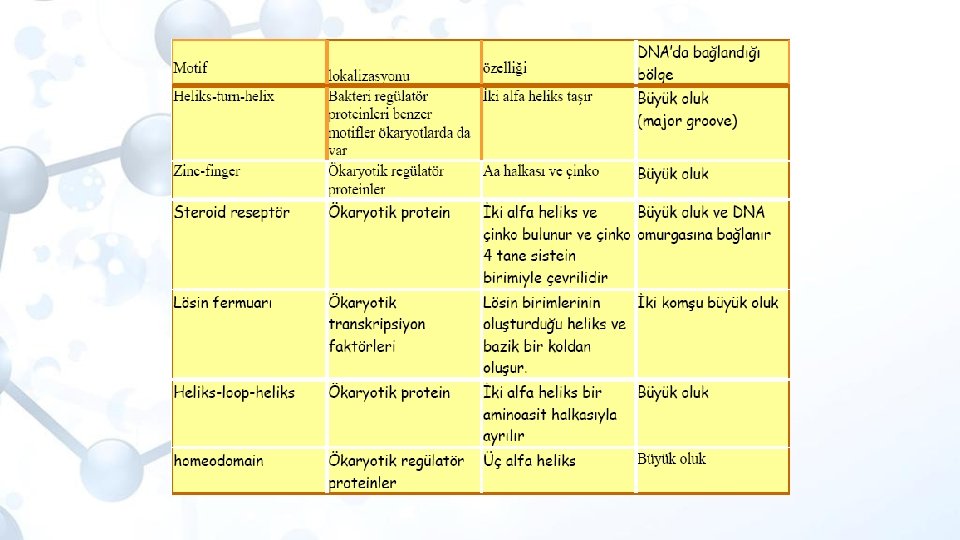
Gerçek aktivatör transkripsiyon faktörleri Ökaryotik transkripsiyon faktörlerinin domainleri çeşitli yapısal şekillerde (motif) bulunur. Sarmal-dönüş-sarmal (helix-turn-helix, HTH) motifi Çinko parmak (zinc finger) Bazik lösin fermuarı (bazic leucine zipper, b. ZIP).
 Motifi Her biri iki alt birim içeren bu proteinlerin DNA’ya iki")
Sarmal-dönüş-sarmal (helix-turn-helix, HTH) Motifi Her biri iki alt birim içeren bu proteinlerin DNA’ya iki alt birimi (dimer) ile bağlandığı ve bağlanma bölgelerinin ikili bir simetri göstermektedir. Bunların üç boyutlu yapıları incelendiğinde, birbirinden oldukça belirgin bir beta (β) dönüsü ile ayrılan farklı iki α–heliksten meydana gelmektedir. Bu yapıya heliks–dönüş–heliks motifi adı verilmiştir. Karboksil uca yakın alt birime Heliks– 3 adı verilmektedir. Heliks– 3, proteinin DNA’ya bağlanmasına yardımcı olduğundan tanıyıcı heliks olarak da adlandırılır.

İki heliks arasındaki mesafe 34 Å dur ve bu mesafe, DNA’nın iki büyük oluğu arasındaki mesafeye eşittir. İki heliks veya altbirim arasındaki polar olmayan etkileşimler bunların oluşturduğu motifi kararlı kılmaktadır. Bu modele göre, heliks– 3’ün alt tarafında bulunan amino asit kökleri, DNA’nın büyük oluğundaki bazlar ile hidrojen bağı oluşturmaktadır. Heliks-2 Tüm ökaryotik organizmalarda hemen evrensel olarak bulunan ve 180 bç uzunluğundaki bir bölgeyi kapsayan homeokutusu (homeobox), HTH yapısı oluşturabilen 60 aminoasitlik homeodomain dizisini belirler. Bu 60 amino asitin çoğu baziktir (arjinin ve lizin). Heliks-3
 motifi Ökaryotik transkripsiyon faktörlerinin yapısal bakımdan ana ailesini oluşturan çinko")
Çinko parmak (zinc finger) motifi Ökaryotik transkripsiyon faktörlerinin yapısal bakımdan ana ailesini oluşturan çinko parmaklar, gen regülasyonuna birçok yönden katılırlar. Her biri belirli bir özgün yapı gösteren çok çeşitli tipte çinko parmak proteinleri vardır. Tipik bir çinko parmak motifinde, iki sistein ve iki histidin amino asitinin belirli aralıklarla tekrarlandığı bir bölge vardır. Bu bölgede Cys–N 2 -4–Cys–N 12 -14–His–N 3–His şeklinde tekrarlayan (N herhangi bir aminoasit) konsensus amino asit dizisi bulunmaktadır. Bu tekrarların herbiri özdeş kısımlarda iki sistein (Cys) ve iki histidin (His) kökü içermekte olup, bu proteinde aynı zamanda dokuz tane de Çinko (Zn) iyonu bulunmaktadır. Herbir çinko atomunun, sisteinlerin kükürt (Cys–S), histidinlerin ise azot (His–N) atomlarına tetrahedral bir şekilde bağlanarak parmak şeklinde yapılar oluşturmaktadır.

Her bir parmak 23 amino asit içerir (Cys ve His amino asitleri arasındaki ilmik bölgesinde yaklaşık 12 -14 amino asit bulunur) ve ilmikler arasındaki bağlantı bölgesi 7 -8 amino asitten oluşur. İlmikte yer alan amino asitler özgül DNA dizileri ile etkileşip ona bağlanırlar. Çinko parmaklar, DNA sarmalının büyük oluğuna bağlanırlar ve DNA’ın yarısına kadar etrafında sarılırlar. Parmak seklindeki yapının ucu, DNA’nın büyük oluğu içindeki bir dizi baz ile, özellikle G’ce zengin zincirdeki bazlarla temas kurar ve hidrojen bağı oluşturur. Transkripsiyon faktöründeki çinko parmak sayısı 2 ile 13 arasında değişir, aynı şekilde, DNA-bağlanma dizisinin uzunluğu da değişkenlik göstermektedir.
 Bu DNA bağlanma domaini, protein-protein dimerinin")
Bazik lösin fermuarı (bazic leucine zipper, b. ZIP) Bu DNA bağlanma domaini, protein-protein dimerinin oluşumuna izin veren lösin fermuar bölgesine bitişiktir. Lösin fermuarında 7 lösin amino asiti birbirlerinden ayrılmıştır ve bu bölgenin iki yanında bazik amino asitler bulunmaktadır. Lösin amino asitlerinin bulunduğu bölge bir sarmal yapı oluşturur ve sarmalların her bir dönüşünde lösin çıkıntıları görülür. Bu şekilde iki molekül bir araya gelince (dimer), lösinler bir “fermuar” gibi birleşir, iki molekül, lösin “fermuarı” ile bir arada tutulur.

Dimer yapısında, fermuar kısma komşu olan iki bazik α-sarmal bölgesi bulunur. Bu bölge, DNA’daki fosfat gruplarına ve özgül bazlara bağlanarak dimerin DNA üzerinde bir makas gibi görünmesine neden olur.

5. Post transkripsiyonel kontrol Advances in analyzing RNA diversity in eukaryotic transcriptomes: peering through the Omics lens [version 1; referees: 3 approved] Sushant Bangru , Auinash Kalsotra

Alternatif splicing-exon shuffling Ekzonların farklı şekilde işlenmesi farklı proteinlerin sentezine neden olur ib. bioninja. com. au alternative splicing.

Alternative splicing
 kuyruğu veya serbest 5 ′")
Halkasal RNA-circ. RNA Circ RNA ların bir poli (A) kuyruğu veya serbest 5 ′ veya 3 ′ ucunun yoktur. Açık 5’ veya 3’ uçlarının olmayışı onları stabilize ederek eksonükleazlara dirençli hale getirir. circ. RNA'ların çoğu, genellikle doku ve gelişim aşamasına özgü bir şekilde ifade edilen genlerin ortasında yer alan ve çok sayıda ekson içeren non coding splice varyantları olduğu gösterilmiştir. Bu varyantların, mikro. RNA aktivitesini antagonize etmede rol oynadığı düşünülmektedir.

RNA editing transkripsiyon sonrası m. RNA’daki bazların değişmesidir. Değişiklikler genellikle, C→U or A→I (I = inozin, G nin kodlama özelliklerine sahiptir) şeklinde olur. Eğer aynı genin bazı RNA ları bu değişikliğe uğrar ise (RNA editing) proteinlerde ekstra çeşitlilik ortaya çıkar.

RNA editing-RNA düzenleme

RNA interferansı, bir m. RNA'nın susturulduğu ve bu şekilde söz konusu m. RNA'dan kodlanmış proteinin inhibe edildiği doğal bir fenomendir. Gerekli m. RNA'yı susturarak istenen proteinin baskılanması için çok etkili bir araç olarak hizmet etmektedir. RNAi mekanizmasına dahil olan temel makro moleküller, iki tip RNA molekülüdür: Mikro RNA (mi. RNA) ve küçük müdahaleci RNA (si. RNA). Bu iki tip RNA, tamamlayıcı bağlanma yoluyla spesifik m. RNA'ya bağlanabilir ve ribozomlar tarafından translasyonu inhibe edebilir.

mi. RNA; kısa kodlamayan RNA nükleotidleridir ve firkete dizilerine sahip olduğundan çift sarmal yapıdadır. Öncül mi. RNA, bir RNase III enzimi ve bir ds. RNA bağlanma alt birimi içeren, mikroişlem birimi adı verilen bir protein kompleksiyle bağlanır; Bu mikroişlem birimi, ön mi. RNA'yı çekirdekten sitoplazmaya, Nükleer gözenek kompleksi (NPC) yoluyla alır ve burada, ön mi. RNA'nın bazı kısımlarını ayıran, ds. RNA'ya özgü bir endonükleaz alt birimi içeren Dicer adlı bir protein aracılığıyla olgun bir mi. RNA ya dönüşür. Eksojen ds. RNA nükleotitleri olan si. RNA molekülleri, dizi bakımından mi. RNA'nunkine benzerdir ancak hücre çekirdeği içinde üretilmezler.

mi. RNA ve si. RNA; bir yolcu ipliği ve bir kılavuz iplik içerir. RNA kaynaklı susturma kompleksi (RISC) adı verilen bir protein ve RNA kompleksi, kılavuz RNA ipliğine bağlanır ve yolcu RNA ipliği bozulur. Bu kılavuz dizisi, belirli bir m. RNA'ya tamamlayıcı olan dizileri içerir. m. RNA'ya bağlanır ve RISC, kılavuz-m. RNA çift ipliğini ayıran m. RNA üzerinde kesikler oluşturur. Kalan m. RNA, sitozolik eksonükleazlar tarafından bozulur. Bu nedenle, m. RNA tarafından kodlanan protein, m. RNA susturulduğundan oluşamaz.
 | Molecular Biology | The Biology Notes")
RNA Interference (RNAi) | Molecular Biology | The Biology Notes

http: //yandex. com. tr/clck/jsredir? from=yandex. com. tr%3 Bgorsel%2 Fsearch%3 Bimages%3 B%3 B&text=&etext=9128. c-Lp. Gx_WJdr 4 h. Eaba. Mv 45 Xz. DIdm. Ink 6 q 3 dgpt. NGFb. A. c 19 a 285152 c 31 b 92 b 87 f 34 c 22 e 5 cf 9 f 5 a 6 da 3 a 59&uuid=&state=iric 5 OQ 0 s. S 1 m. Pitaa 3 mx. JE 61 AVKS 1 Y 9 si. PMm. VFs. WPIWEtr. Eg. Mmapww, , &data=e. Ewy. M 2 l. DYU 9 Gd 1 Vt. OEow. Yzd. MS 3 hk. WWZSU 3 F 5 ZVFQ a. Uot. NFhq. V 2 REd. Fd. GOWJaan. Nh. Wk. Zlb. Uh. UV 2 d. Gclgz. M 1 l. Ca. XBhcmp 6 ZUs 4 WXRENE 9 USnpl. NHJo. RWRYd. VZIV 1 E 4 e. Xp. PMGt 5 a 1 Ay. VEt. Md 3 F 1 Skd. ITj. Jl. UU 1 Kb. Dd. Ob 2 VJd. Flo. SGh 5 Rj. NYZThu. NFp. SOW 1 Hd. WVBTHh. PVGFBLCw, &sign=37 eed 241 e 4 c 8 e 8 bf 2047 f 11727 b 63040&keyno=IMGS_0&b 64 e=2&l 10 n=tr

Protein modifikasyonudegradasyonu Protein modifikasyonu katlanma, kesilme, şeker gruplarının eklenmesi, taşınma Protein degradasyonu Ubikutin ekleme Proteazomda parçalanma

https: //www. slideshare. net/jayswan/chapter-18 -generegulation

KAYNAKLAR • Genomlar 3 T. A. BROWN. 3. baskıdan çeviri, Çeviri editörleri: Fevzi BARDAKÇI-Celal ÜLGER, NOBEL • Modern Moleküler Biyoloji Prof. Dr. Nihat DİLSİZ, Palme Yayıncılık, 2017 • Moleküler Hücre Biyolojisi, Prof. Dr. Hasan Veysi GÜNEŞ , 2014, istanbul Tıp Kitabevi • Moleküler Hücre Biyolojisi, 6. baskıdan çeviri, 2011, Palme Yayıncılık, Çeviri editörleri: Prof. Dr. Hikmet Geçkil, Prof. Dr. Murat Özmen, Prof. Dr. Özfer Yeşilada • Temel Moleküler Biyoloji, 2. baskıdan çeviri, Çeviri Editörü: Prof. Dr. Ali Osman Beldüz, 2014 78
- Slides: 78