Mikrobiyal insektisitler Bacillus thuringiensis Zararl kimyasal pestisitlerin yerine

Mikrobiyal insektisitler: Bacillus thuringiensis

Zararlı kimyasal pestisitlerin yerine çevre dostu mikrobiyal böcek öldürücülerin kullanılması, yıkıcı mahsul zararlılarının kitlesel kontrolünde önemli bir alternatiftir. Giriş Biyokontrol ajanları (makro ve mikro) için küresel pazar, yıllık % 16 büyüme ile yaklaşık 3, 5 milyar USD'dir ve bu, küresel pestisit ticaretinin yaklaşık % 8'ini (50 milyar USD) oluşturmaktadır. Mikrobiyal insektisitlerin payı yaklaşık 807 milyon USD'dir (BCC Araştırma Raporu 2015; Lacey ve diğerleri 2015; Velivelli ve diğerleri 2014).
, parasporal kristal proteinleri veya endotoksinleri (Cry) üreten aerobik, spor oluşturan,")
Bacillus thuringiensis (Bt), parasporal kristal proteinleri veya endotoksinleri (Cry) üreten aerobik, spor oluşturan, gram pozitif ve entomopatojenik bir bakteridir. Bu Cry proteinleri, Lepidoptera, Coleoptera ve Diptera gibi çok çeşitli böcek zararlıları için toksiktir. Bt, geçen yüzyılda en başarılı biyoinsektisit olarak kabul edilmiştir. Giriş Şu anda 98'den fazla (424 milyon ABD doları) formüle edilmiş püskürtülebilir bakteriyel pestisitten oluşmaktadır (Lacey ve diğerleri 2015). Bt türü genellikle, farklı filogenetik ve serotipleme özelliklerine sahip farklı alt türlerde kategorize edilen büyük bir farklı alt tür ailesinden oluşur (Bt subsp. kurstaki, Bt subsp. aizawai, Bt subsp. tenebrionis, Bt subsp. israelensis, vb. ). Ek olarak, her Bt alt türü farklı suşlardan ve serotiplerden oluşur (Seifinejad ve diğerleri 2008).

Bt, hızlı etkili ve konakçıya özgü bir biyoinsektisit olarak bilinir, bu nedenle hedef olmayan organizmalar üzerindeki olumsuz etkileri çok sınırlıdır. Giriş Dahası, üretimi (yukarı ve aşağı akış süreçleri) ve uygulaması (geleneksel püskürtme veya genetiği değiştirilmiş (GM) Bt mahsulleri) kolay ve ucuzdur (Jain ve diğerleri 2016; Lacey ve diğerleri 2015). Buna göre Bt, transgenik bitkileri farklı zararlılara karşı dirençli hale getirmek için bitki genetik mühendisliğinde cry genlerinin kaynağı olarak etkin bir şekilde kullanılmıştır. Ayrıca bitki patojenik nematodlarını kontrol etmek için bir nematisit olarak kullanılma potansiyeline sahiptir (Iatsenko ve diğerleri 2014 a, b, Salehi Jouzani ve diğerleri 2008 b).

Son araştırmalar farklı Bt suşlarının daha yeni potansiyelleri olduğunu göstermiştir. Bu yeni özellikler arasında; Bitki büyümesini teşvik etme (Armada ve diğerleri 2015 a, b), Giriş Ağır metallerin ve diğer kimyasalların biyoremediasyonu (Aceves. Diez ve diğerleri 2015; Dash ve diğerleri 2014; Melo ve diğerleri 2016), Antikanser faaliyetleri ( Periyasamy ve diğerleri 2016), Polimer üretimi (Singh ve diğerleri 2013) ve Bitki ve hayvan patojenik mikroorganizmalara karşı antagonistik etkiler (Gutiérrez-Chávez ve diğerleri 2016; Roy ve diğerleri, 2013). verilebilir.

Bt nin etki alanları

Bt genom çalışmalarında son gelişmeler Yeni nesil dizilemede son gelişmeler ve yeni genomik, transkriptomik, proteomik ve transkriptomik gibi -omik teknolojileri, Bacillus türleri arasındaki ve ayrıca farklı Bt alt türleri ve suşları arasındaki genom çeşitliliğine ilişkin önemli veriler sağlamıştır. Bt genom projelerinin yeni patojenik genlerin ve ilgili düzenleyici faktörlerin tespitini artırması ve hızlandırması beklenmektedir. Ayrıca, farklı özelliklere ve aktivitelere sahip Bt toksin proteinlerini incelemek için genomik, transkriptomik, proteomik ve metabolomiklerin bir kombinasyonu kullanılabilir (Dong ve ark. 2016).

Şimdiye kadar, 60 tan fazla Bt suşuna ait tam ve parçalı genom dizileri (yaklaşık 30 tam sekans) Gene. Bank'a gönderilmiştir. Bt genom çalışmalarında son gelişmeler İncelenen Bt suşlarının tam uzunluktaki genomu (bir ila birden fazla plazmit dahil) 5, 3 ila 6, 87 Mb arasındadır. Çalışılan Bt suşlarındaki gen sayısı 5343 ile 7227 arasında ve plazmit sayısı 1 ile 13 arasında değişmektedir. Bt genomlarının guanin-sitozin içeriği (GC)% 31. 4 ile% 35. 48 arasındadır (Tablo 1). Bu buldular, incelenen suşlar arasındaki geniş genetik çeşitliliği doğrulamaktadır. Dolayısıyla, yeni suşların keşfedilmesi, büyük olasılıkla yeni toksin genleri ve proteinlerin tespitini mümkün kılacaktır.
")
Bt insektisidal genler ve konak özgüllüğü Bt flagellar antijenlere göre 72 antijenik grup (serotip) ayırt edilmiştir (Blackburn ve ark. 2013; Lecadet ve ark. 1999; Lecadet 2013). Crickmore vd. (2016), Bt toksinleri için, sürekli olarak güncellenen konakçı böcekler hakkındaki bilgilere bağlantılar içeren özel bir veritabanı tasarlamıştır (www. lifesci. sussex. ac. uk/Home/Neil_Crickmore/Bt/). Bu veri tabanındaki (Haziran 2016) en son güncellenen verilere dayanarak, dünyanın farklı bölgelerinden izole edilen Bt suşlarında farklı entomopatojenik proteinli toksinleri kodlayan yaklaşık 952 toksin geni tanımlanmış ve karakterize edilmiştir.

Bu toksinlerin çoğu sporülasyon aşamasında üretilen parasporal kristallerdir. Bt insektisidal genler ve konak özgüllüğü Parasporal inklüzyon cisimcikleri, delta-endotoksinler olarak bilinen kristalin proteinler içerir ve Cry ve Cyt proteinleri olmak üzere iki aileye ayrılır. Amino asit sekans benzerliklerine dayanarak, şimdiye kadar 770 farklı cry genli 74 cry geni ailesi (cry 1 – cry 74) ve 38 cyt geninden oluşan üç cyt ailesi (cyt 1 -cyt 3) karakterize edilmiştir. Diğer böcek öldürücü (insektiisidal) proteinler, büyümenin vejetatif fazı sırasında üretilen bitkisel böcek öldürücü proteinlerdir (Vips). Şimdiye kadar, dört gruba (vip 1– vip 4) ayrılan yaklaşık 138 farklı vip geni tanımlanmış ve karakterize edilmiştir (Tablo 2).

Bt genom çalışmalarında son gelişmeler Cry, Cyt ve Vip protein içeriklerine bağlı olarak, her bir suş özellikle lepidopteran, dipteran, coleopteran veya hymenopteran zararlılarına ve hatta akarlar ve nematodlar gibi diğer omurgasızlara karşı aktif olabilir (Abdelmalek ve diğerleri 2015; Salehi Jouzani ve diğerleri. 2008 a, b). Cry 1 ailesi, 275 cry geni içeren 14 alt aile (cry 1 A – N) içerir. cry 1 genlerinin çoğu lepidopteran zararlılarına karşı etkindir. Bu aileden gelen cry 1 b ve cry 1 I genleri aynı zamanda koleopteran zararlılarına karşı da aktiftir (Nazarian ve diğerleri 2009). Cry 2 ailesi yaklaşık 82 gen ile ikinci sırada yer alır ve aktiviteleri çoğunlukla lepidopteran veya dipteran zararlılarına karşıdır. Cry 3 familyası, çoğunluğu koleopteran böceklere karşı aktif olan 19 gen içerir (Tablo 2).

cry 1, cry 9, cry 15, cry 20, cry 51, cry 54, cry 59 ve vip genleri lepidopteran zararlılarına karşı esas olarak etkindir. cry 2, cry 4, cry 10, cry 11, cry 16, cry 17, cry 19, cry 24, cry 25, cry 27, cry 29, cry 30, cry 32, cry 39, cry 40, cry 44, cry 47, cry 48, cry 49, cry 52 ve cyt genleri dipteran zararlılara karşı etkindir. Bt genom çalışmalarında son gelişmeler cry 3, cry 7, cry 8, cry 14, cry 18, cry 22, cry 23, cry 26, cry 28, cry 34, cry 35, cry 366, cry 37, cry 38, cry 43 ve cry 55 ise coleopteran spesifik genlerdir (Tablo 2). Ayrıca, son yirmi yılda cry 5, cry 6, cry 12, cry 13, cry 14, cry 21 ve cry 55 gibi bazı cry genlerinin bitki ve hayvan nematodlarına karşı toksisitesi olduğu kanıtlanmıştır (Ruan ve diğerleri 2015; Salehi Jouzani et al. 2008 b). Bununla birlikte, Cry proteinlerinin yüksek konsantrasyonları kullanıldığında bildirilen bazı nematisidal aktivitelerin gözlemlendiği dikkate alınmalıdır.

Son yıllarda, modern biyoteknoloji uygulamasında, özellikle tarımda hızlı ilerlemelere tanık olunmuştur. Bt ve bitki genetik mühendisliği Dünya genelinde GDO'lu mahsullerin küresel hektarı, sosyoekonomik ve çevresel avantajları nedeniyle son 20 yılda önemli ölçüde artmış ve 2015 yılında 179, 7 milyon hektara ulaşmıştır (Salehi Jouzani ve diğerleri 2008 c; Salehi Jouzani 2012; Tohidfar ve Salehi Jouzani 2008; James 2015). Bitki genetik mühendisliğinde en yaygın olarak kullanılan özellikler herbisit ve zararlılara karşı dayanıklılıktır. Bitkilerdeki zararlılara karşı direnci artırmak için Bt toksin genleri yaygın olarak kullanılmıştır. 2015 yılında, Bt transgenik mahsullerin alanı yaklaşık 75 milyon hektardır (58, 5 milyon hektar yığın Bt / herbisit toleransı ve 18 milyon hektar Bt mahsuller). Bu GM mahsulleri, lepidopteran ve / veya koleopteran zararlılarına karşı direnç için bir veya daha fazla farklı cry geni içerir (James 2015). Bt mahsulleri, 1996 -2014 boyunca pestisit uygulamasında 583 milyon kg'dan fazla azalma sağlamıştır (Brookes ve Barfoot 2015; James 2015).

• 1996'dan beri, mısır, pamuk, patates, soya fasulyesi, domates, kavak, çeltik ve patlıcan dahil olmak üzere sekiz bitkinin 198 Bt GM çeşitleri ve hatları ticari sürüm için onaylanmıştır (Şekil 2). Bt ve bitki genetik mühendisliği

Bt ve bitki genetik mühendisliği Sırasıyla, 115, 42 ve 30 çeşit ve hatlı mısır, pamuk ve patates en çok onaylanan Bt GM mahsulleridir (ISAAA’nın GM Onay Veritabanı 2016). Cry 1 Ab, cry 1 A. 105, cry 1 Ac, cry 1 F, cry 2 Ab, cry 2 Ae ve vip 3 A dahil yedi antilepidopteran cry ve vip geni lepidopteran genlerine direnci artırmak için kullanılmıştır. Cry 1 Ab, cry 1 F ve cry 1 Ac sırasıyla 61, 51 ve 32 GM çeşitlerinde kullanılan lepidopteran dirençli mahsuller üretmek için en çok kullanılan genlerdir (Şekil 3).

Bt ve bitki genetik mühendisliği
 içerir.")
Bazı Bt mahsulleri birden fazla cry veya vip geni (iki veya üç) içerir. Bu gen piramit sistemleri, transgenik bitkilerde üretilen Bt toksinlerine karşı potansiyel haşere direncini geciktirmek için geliştirilmiştir. Bt ve bitki genetik mühendisliği Antikoleopteran genleri içeren onaylanmış Bt çeşitlerinin sayısı yaklaşık 111'dir, bunların bazıları ayrıca anti-lepidopteran zararlıları da içerir. cry 3 Aa, cry 3 B, cry 34 Ab 1 ve cry 35 Ab 1 dahil olmak üzere dört anticoleopteran cry geni, koleopteran haşerelere karşı direnci arttırmak için kullanılmıştır. cry 34 Ab 1 ve cry 35 Ab 1 bir hibrit gen olarak kullanılmıştır. Coleopteran zararlılarının dirençli mahsullerini üretmek için en çok kullanılan genler olan cry 3 A ve cry 34 Ab 1 – cry 35 Ab 1, sırasıyla 60 ve 34 GM çeşitlerinde kullanılmıştır (Şekil 4).

Bt ve bitki genetik mühendisliği
, ve kök-düğüm nematodları (Meloidogyne spp. )")
Kist nematodları (Heterodera ve Globodera spp. ), ve kök-düğüm nematodları (Meloidogyne spp. ) dahil bitki parazitik nematodları delici / emici zararlı böcekler olup farklı mahsullere şiddetli zarar verirler. Biyolojik nematosit olarak Bt Bu nematodlar yıllık küresel olarak yaklaşık 125 milyar dolarlık verim zararına neden olur (Chitwood 2003; Yu et al. 2014, 2015; Zhang vd. 2012). Ayrıca, hayvan paraziti nematodları, veterinerlik hizmetlerinin maliyetini artırarak, hayvan büyümesini geciktirerek ve hatta ölüme neden olarak hayvansal üretimi engelleyen en önemli faktörlerden biri olarak bilinmektedir (Sinott ve ark. 2012). Kimyasal nematisitler, kök-düğüm nematodlarını kontrol etmenin en güncel yolu olmaya devam etse de, çevre güvenliği ve halk sağlığı ile ilgili artan endişeler, bu tür nematisitlerin geri çekilmesine veya kullanımının kısıtlanmasına yol açmaktadır (Yu ve ark. 2015).

Biyolojik nematosit olarak Bt Bazı Bt türleri, nematodların sindirim sistemi içinde enfekte edebilir, filizlenebilir ve çoğalabilir (Ruan ve diğerleri 2015). Cry 5, cry 6, cry 12, cry 13, cry 14, cry 21 ve cry 55 gibi bir veya daha fazla kristal protein ailesi içeren Bt suşlarının nematisidal aktivitelere sahip olduğu belgelenmiştir (Guo ve diğerleri 2008; Luo ve diğerleri 2013 a, b ; Salehi Jouzani ve diğerleri 2008 b; Yu ve diğerleri 2015; Zhang ve diğerleri 2012) (Şekil 1 ve 5). Ayrıca, bu Cry proteinleri, Bt suşlarında mevcut olduklarında nematodlar üzerinde sinerjistik etkilere sahiptir (Yu ve diğerleri 2014). Buna göre, rekombinant nematod aktif Cry proteinlerinin ifadesi bitkilerde bitki endoparazitik nematodlara karşı koruma sağlar (Li ve ark. 2007, 2008).
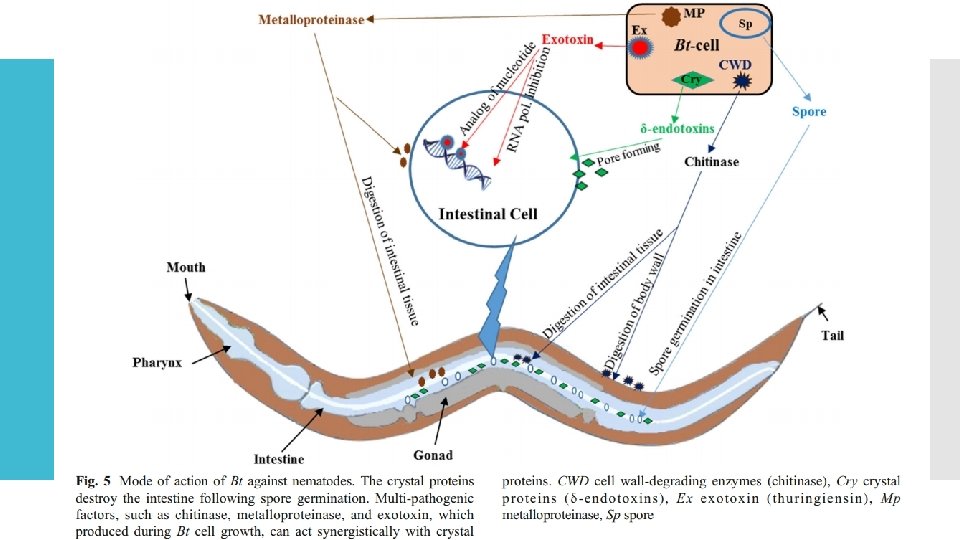

Thuringiensin, kitinaz ve metaloproteinaz gibi diğer birkaç Bt bileşiği nematisidal aktiviteler göstermektedir (Şekil 5). Biyolojik nematosit olarak Bt Lantibiyotikler, enterotoksinler, hemolizinler ve çoğunlukla transkripsiyonel aktivatör Plc. R tarafından kontrol edilen proteazlar dahil olmak üzere nematisidal faktörleri kodlayan diğer genler de doğrulanmıştır (Ruan ve diğerleri 2015; Zhou ve diğerleri 2014). Peng vd. (2016), Cry 5 ve Cry 6 proteinlerinin nematisidal aktivitelerini arttırmak için metaloproteinaz Col. B (kolajenaz proteini) varlığının çok gerekli olduğunu kanıtlamıştır. Ruan vd. (2015), nematodlarla Bt etkileşimleri için diğer iki alternatif mekanizma (nekrotrofizm ve fosz) önermiştir.

Biyolojik nematosit olarak Bt Son dizileme projeleri, nekrotrofik yaşam evresinde yer alan genlerin Npr. R (aktivitesi Npr. X sinyal peptidine bağlı olan ve nekrotrofizm mekanizmasına dahil olan bir transkripsiyonel faktör) regülatörünün kontrolü altında olduğunu doğrulamıştır (Dubois ve diğerleri 2012). Üç protein, keratinolitik proteinaz, kollajenaz (pleiotropik transkripsiyonel faktör (Plc. R) ile düzenlenir) ve immün inhibitör A, Bt'nin nekromenik yaşam tarzını mümkün kılar. Keratinolitik proteinaz, nematod kütikülündeki kolajen içeriğini sindirir. İkinci alternatif mekanizma, Bt'nin nematod tarafından ya yüzeyinde ya da bağırsak yolunda konağı öldürmeden taşındığı foresidir (Ruan ve ark. 2015).

Bt suşlarının nematisidal aktiviteleri, Serbest yaşayan nematodlar Biyolojik nematosit olarak Bt Caenorhabdita elegans, Pristionchus pacificus ve Chiloplacus tenuis Hayvan parazitik nematodları Ascaris suum, Distolabrellus veechi, Haemonchus contortus, Trichostrongylus sp. ve Ostertagia circincta Bitki parazitik nematodlar Meloidogyne incognita, Meloidogyne halpa, Pratylenchus, Tylenchorhynchus sp. , Ditylenchus destructor ve Aphelenchoides sp. üzerinde test edilmiştir.

Bazı akar ve kene türleri, insanları ve hayvanları doğrudan kolonize eder ve ayrıca hastalığın bulaşması için vektör görevi görür veya alerjenik hastalıklara neden olur. Bt'nin akarlar üzerindeki etkisine ilişkin bilgiler nadir olmakla birlikte, birkaç in vitro ve in vivo çalışmada bazı Bt suşlarının akarisidal aktivitesi bildirilmiştir (Erban ve diğerleri 2009; Dunstand-Guzmán ve diğerleri 2015; Alquisira. Ramírez ve diğerleri. 2014). Bt lerin akarisidal etkileri İlk raporlarda Hassanain ve ark. (1997) tarafından, yumuşak kene Argas persicus ve sert kene Hyalomma dromedarii'ye karşı üç Bt alt türünün (kurstaki, israelensis ve thuringiensis) akarisit aktiviteleri değerlendirilmiştir. Bt. var. Kurstaki ve Bt var. israelensis sırasıyla en yüksek toksisiteyi göstermiştir. Bt var. kurstaki nin akarisit etkisi çeşitli hayvan ve insan hastalıkları için vektör görevi gören siyah bacaklı keneye karşı (Ixodes scapularis Say) da doğrulanmıştır (Zhioua ve diğerleri 1999). Başka bir çalışmada, Cry 3 A toksini üreten Bt var. tenebrionis, Acarus siro L. , Tyrophagus putrescentiae, Dermatophagoides farinae ve Lepidoglyphus destructor akarlarına karşı yüksek toksisite (LC 50 25 ila 38 mg / g) göstermiştir (Erban ve ark. 2009). Alquisira-Ramírez ve diğerleri. (2014), arı Apis mellifera'nın (Hymenoptera: Apidae) hemolimfini besleyen ektoparazitik bir akar olan Varroa destroyer (Acari: Varroidae) 'den yüksek akarisidal aktiviteye sahip bazı Bt suşlarını izole etmiş ve karakterize etmiştir

Başka bir grup, ilk olarak Bt suşu GP 532'nin tavşan kulağının ortak bir ektoparaziti olarak bilinen Psoroptes cuniculi akarı üzerindeki akarisit aktivitesini (LC 50 1. 3 mg / ml ve LT 50 68 h) bildirmiştir (Dunstand-Guzmán ve diğerleri 2015). Bt lerin akarisidal etkileri Son zamanlarda, farklı ekinleri ve süs bitkilerini istila eden bir emici akar olan Tetranychus macfarlanei'ye (LC 50 8 mg / ml) etkili bir şekilde toksik olan Malabari keçisinin rumeninden yeni bir Bt suşu (BPU 5) izole edildi (Neethu ve diğerleri 2016). Ahmed vd. (2016), Bt var. israelensis (% 81. 22 mortalite) ve tenebrionis (% 90. 91 mortalite) in, depolanan gıda ürünlerinin kozmopolit bir zararlısı olan küf akarı T. putrescentiae'ye (Schrank) karşı 4 hafta sonra 32 mg / kg oranında akarisit etkisini rapor etmiştir.

Bt suşlarının farklı kenelere ve akarlara karşı akarisidal aktivitesinin doğrulanmasına rağmen, akarisidal Bt suşlarının etki mekanizması henüz bilinmemektedir. Bt lerin akarisidal etkileri Bununla birlikte, incelenen akarların sindirim sistemi üzerinde tripsin, alkalin fosfataz ve bazı aminopeptidazlar gibi enzimlerin varlığı, akarların bağırsak hücrelerindeki değişikliklerin Bt protoksinlerinin aktivasyonundan kaynaklanabileceğini düşündürmektedir (Dunstand-Guzmán ve ark. 2015 ). Akar sindirim sistemindeki Bt'nin etki mekanizmasının araştırılması, Bt ile ilgili gelecekteki çalışmalarda dikkate alınması gereken konulardan biridir. Bununla birlikte, yüksek akarisidal aktiviteye sahip olmak için, Bt'nin ticari düzeyde biyoakarit olarak kullanılmasını pratik olmayan kılan yüksek doz Cry proteinleri / sporları (mg / ml) gereklidir. Şu anda dünyada akar ve kene türlerinin kontrolü için piyasada satılan Bt ürünü bulunmamaktadır. Bu nedenle, gelecekte daha güçlü akarisidal aktiviteye sahip yeni Bt suşlarının araştırılması gerekli olacaktır.

Bitki büyümesini destekleyen bakteri olarak Bt Genellikle, bitki büyümesi ve gelişmesi üzerinde yararlı etkileri olan bakteri türleri, bitki büyümesini teşvik edici rizobakteriler olarak adlandırılır (Mishra ve diğerleri 2009 a). Bazı Bt türleri bitki köklerinde kolonize olurlar ve bitki büyümesini teşvik edici özelliklere sahiptir. Bu Bt suşları, tarımda biyogübre olarak, tek başına veya diğerleriyle karışım halinde kullanılma potansiyeline sahiptir. Bai vd. (2003), Bt sp. NEB 17'nin, Bradyrhizobium japonicum ile soya fasulyesi bitkileri üzerine birlikte aşılandıklarında, Bacillus subtilis suşlarına kıyasla soya fasulyesi nodülasyonunu, büyümesini ve verim parametrelerini önemli ölçüde artırdığını doğrulamıştır. Rhizobium leguminosarum-PR 1 ile IAA üreten bir Bt suşu olan KR 1'in eşokülasyonu, yalnızca R. leguminosarum-PR 1'in aşılanmasına kıyasla tarla bezelyesi ve mercimek büyümesini önemli ölçüde teşvik edebilmektedir (Mishra ve diğerleri, 2009 a).

Bitki büyümesini destekleyen bakteri olarak Bt Bt-KR 1'in B. japonicum-SB 1 ile kookülasyonu da soya fasulyesi bitkilerinin büyümesini teşvik etmiştir ve rizobiyal aşılama ve kontrol ile karşılaştırıldığında nodül sayısı, sürgün ağırlığı, kök hacmi ve toplam biyokütlede önemli bir artış sağlanmıştır (Mishra ve ark. . 2009 b). Birçok Bt suşu, abiyotik stres koşullarında bitki büyümesini artıran bazı metabolitler üretir. Bu bileşikler, ACC deaminaz, indol-3 asetik asit (IAA), prolin ve fosfat çözündürme enzimlerini içerir (Şekil 6). Armada vd. (2015 a), Bt'nin tek başına veya arbusküler mikorizal mantarlarla (AMF) karıştırılması durumunda, bitki sürgünlerinde sürgün büyümesi, biyokütle (% 20'den fazla) ve mikro besin elementlerinde önemli bir artışa neden olabileceğini göstermiştir.

Bitki büyümesini destekleyen bakteri olarak Bt Ayrıca, antioksidan enzim aktivitelerini artırarak (süperoksit dismutaz, katalaz ve askorbat peroksidaz) ve lipitlerin bitki oksidatif hasarını (malondialdehit) azaltarak oksidatif stresi önemli ölçüde azaltabilir. Başka bir çalışmada, kuraklık stresi altında mısıra Bt ve AMF'nin kombine aşılanması, bitkide besin birikimini önemli ölçüde artırabilir ve lipidlere oksidatif hasarı ve prolin birikimini azaltabilir (Armada ve diğerleri 2015 b). Ayrıca, otokton mikroorganizmaların (bir Bt ve AMF konsorsiyumu) uygulanması, artan besin içeriği ve nispi su içeriği ve azalan stoma iletkenliği, elektrolit sızıntısı, prolin ve askorbat peroksidaz aktivitesi yoluyla kuraklık koşullarında doğal bir kurak toprakta Trifolium repens için su stresinin azalmasında önemli bir artış sağlamıştır. (Ortiz ve ark. 2015).
, Bt sp. NEB 17'den saflaştırılmış")
Bitki büyümesini destekleyen bakteri olarak Bt Lee vd. (2009), Bt sp. NEB 17'den saflaştırılmış bakteriyosinin (thuricin-17) yapraklara (sprey) veya köklere (ıslatma) uygulanmasının hem bir C 3 dikot (soya fasulyesi) hem de bir C 4 monokot (mısır) bitkisinin büyümesini doğrudan uyardığını doğrulamıştır. Thuricin-17'nin N 2 sabitleyici B. japonicum ile su stresi koşullarında uygulanması bitki biyokütlesini (% 17), kök biyokütlesini (% 37), nodül biyokütlesini (% 55), kök absisik asidi (% 30) ve toplam nitrojen miktarını (% 17) önemli ölçüde artırabilir (Prudent ve ark. 2015). Son zamanlarda, Cherif-Silini ve ark. (2016), Cezayir'in farklı bölgelerinde buğday rizosferinden izole edilen Bt ve B. subtilis suşları için bitki büyümesini teşvik eden rizobakteri (PGPR) aktivitesini bildirmişlerdir. Bu suşlar, maksimum biyo gübreleme (fosfat çözündürme), biyostimülasyon (IAA üretimi) ve biyokontrol aktiviteleri (siyanhidrik asit, sideroforlar ve 2, 3 -butandiol üretimi ve antifungal aktivite) göstermiştir. Bt'nin olası PGPR mekanizmaları Şekil 6'da özetlenmiştir.

Bitki büyümesini destekleyen bakteri olarak Bt

Bitki büyümesini destekleyen bakteri olarak Bt suşlarının PGPR aktivitesi ile ilgili önceki çalışmaların sonuçları çok umut vericidir. Bununla birlikte, şu anda, biyo gübre pazarında ticari olarak temin edilebilen Bt bazlı PGPR formülasyonu bulunmamaktadır. Güçlü PGPR aktivitelerine sahip yeni Bt suşları bularak ve bu Bt suşlarının PGPR aktivitesinin ayrıntılarını keşfederek, PGPR Bt suşlarının ticari üretimi yakın gelecekte farklı ekin sistemleri için mevcut olacaktır.

Bitki ve insan patojenik fungus antagonisti olarak Bt Genel olarak, biyokontrol ajanlarının antifungal etkileri, antibiyotikler, lipopeptitler, sideroforlar, uçucu organik bileşikler, ikincil metabolitler ve hücre duvarını parçalayan enzimler gibi çeşitli antifungal bileşiklerden dolayıdır. Bitkilerde sistemik direnci indükleyen sinyal molekülleri dikkate alınmalıdır (Gao ve diğerleri 2014; Pane ve diğerleri 2012; Shrestha ve diğerleri 2015). Bt tarafından sentezlenen Cry proteinleri, herhangi bir antifungal aktivite göstermezler. Bununla birlikte, bazı Bt suşları, hücre duvarını parçalayan enzimler, lipopeptid fengisin, uçucu bileşikler (VOC'ler) ve sistemik direnci indükleyen sinyal molekülleri dahil olmak üzere antifungal bileşikler üretir (Şekil 6). Fusarium, Sclerotium, Colletotrichum, Rhizoctonia ve Botrytis gibi farklı bitki patojenik funguslara karşı Bt suşlarının antifungal aktiviteleri daha önce gösterilmiştir (Akram ve diğerleri 2013; Reyes-Ramírez ve diğerleri 2004; Sadfi ve diğerleri 2001; Shrestha ve diğerleri 2015; Tang ve diğerleri 2012; Zheng ve diğerleri 2013).

Kitinaz aktivitesi, Bt suşlarında tespit edilen en önemli antifungal ajanlardan biri olarak bilinir. Kitinaz üreten Bt suşları, Bitki ve insan patojenik fungus antagonisti olarak Bt Fusarium roseum var. Sambucinum (patates yumrularının kuru çürümesinin nedensel ajanı ) (Sadfi ve ark. 2001), Sclerotium rolfsii Sacc (Reyes-Ramírez ve ark. 2004), Penisillum chrysogenum (insan hastalığının nedensel ajanı), Rhizoctonia sp. ve Fusarium oxysporum (Gomaa 2012), Sclerotinia minor ve Sclerotinia sclerotiorum, (marul damlasının nedensel ajanları ) (Shrestha ve diğerleri 2015), Urocystis tritici buğday bayrağı isi (Tao ve diğerleri 2014), Fusarium verticillioides (mısır patojeni) (Rocha ve diğerleri 2014) varlığına karşı yüksek antifungal aktiviteler göstermiştir.

Bitki ve insan patojenik fungus antagonisti olarak Bt Ayrıca, son çalışmalar, bitkilerde farklı fungus patojenlerine karşı Bt suşlarının sistemik direnç indüksiyonunu doğrulamıştır. Örneğin, 2 haftalık domates fidelerinin kökleri Bt-199 (CFU 103) vejetatif hücreleriyle 30 dakika aşı içinde tutularak hazırlandığında ve daha sonra saksılara aktarıldığında, bakteri domatesde F. oxysporum lycopersici solgunluğuna karşı sistemik direnç oluşturabilir. Bu direnç, toplam fenolik (1, 7 kat) ve polifenol oksidaz (1. 3 kat), fenil amonyak liyaz (1. 8 kat) ve peroksidaz (1. 4 kat) dahil olmak üzere savunmayla ilgili enzimlerin miktarında önemli bir artışla sağlanır. Bununla birlikte, bu metabolitlerin Bt suşları tarafından nasıl artırıldığına dair mekanizma net değildir (Akram ve ark. 2013).

Bitki ve insan patojenik fungus antagonisti olarak Bt Başka bir çalışmada, Bt-H 3'ten ekstrakte edilen kitinaz, Pyricularia grisea (% 72, 2), Thantephorus cucumris (Rhizoctonia solani) (% 62, 6), Fusarium vasinfectum (% 44, 6), Fusarium gramineum (%50, 0) ve F. oxysporum (% 55. 8) dahil olmak üzere çeşitli patojenik fungusların miselyal büyümesini önemli ölçüde inhibe edebilir. Tür, fenilalanin amonyak liyaz (PAL) ve peroksidaz (POD) dahil çeltik fidelerinin savunma enzim aktivitesini önemli ölçüde artırabilir (Tang ve ark. 2012).

Bitki ve insan patojenik fungus antagonisti olarak Bt Başka bir çalışmada, Bt-H 3'ten ekstrakte edilen kitinaz, Pyricularia grisea (% 72, 2), Thantephorus cucumris (Rhizoctonia solani) (% 62, 6), Fusarium vasinfectum (% 44, 6), Fusarium gramineum (%50, 0) ve F. oxysporum (% 55. 8) dahil olmak üzere çeşitli patojenik fungusların miselyal büyümesini önemli ölçüde inhibe edebilir. Tür, fenilalanin amonyak liyaz (PAL) ve peroksidaz (POD) dahil çeltik fidelerinin savunma enzim aktivitesini önemli ölçüde artırabilir (Tang ve ark. 2012).

Bitki ve insan patojenik fungus antagonisti olarak Bt Bt'nin diğer bir antifungal mekanizması, fengisin benzeri ve uçucu bileşiklerin üretimidir (Şekil 6). Kim vd. (2004), Bt CMB 26'dan fitopatojenik antraknoz mantarı Colletotrichum gloeosporioides, Escherichia coli ve lahana beyazı kelebeğe (Pieris rapae Crucivora) karşı güçlü toksisiteye sahip bir lipopeptidi (fengisin) saflaştırmıştır. Başka bir çalışma, Bt-TB 72'nin 2 -nonanon, β-benzenetanamin, 2 dekanon ve timol gibi farklı uçucu bileşikler ürettiği bildirilmiştir. Bu bileşikler, hasat sonrası mangolarda C. gloeosporioides'in misel büyümesini, sırasıyla in vitro ve in vivo düzeylerde, % 80. 07 ve 87. 06 oranında inhibe edebilir (Zheng ve ark. 2013).

Bitki ve insan patojenik fungus antagonisti olarak Bt Birçok Bt suşu, Candida albicans, Aspergilus niger (Roy ve diğerleri 2013) ve P. chrysogenum (Gomaa 2012) gibi bazı insan ve hayvan patojenik funguslarını da kontrol edebilir. Örneğin, Bt suşu SM 1, C. albicans ve A. niger'e karşı yüksek antifungal aktiviteye sahip fengisin benzeri bir lipopeptit üretir (Roy ve diğerleri, 2013). Bt suşlarının insan ve hayvan patojenik mantarlara karşı antagonistik etkileri hakkında bilgi daha azdır ve buna bağlı olarak, bu antifungal özellikleri bitki koruma, ilaç ve gıda endüstrilerinde kullanmanın yollarını bulmak için gelecekte daha detaylı araştırmalar yapılması gerekmektedir.

Bazı Bt suşları, bitki ve insan patojenik bakterilere ve gıda bozunmasında rol oynayan bakterilere karşı antibakteriyel aktivitelere sahip olabilir. Genellikle prokaryotlar, diğer mikroorganizmalara karşı savunmalarını güçlendirmek için farklı antimikrobiyal peptitler üretir. Patojenik bakterilerin antagonisti olarak Bt Bakteriyosinler, 3 ila 12 k. Da arasındaki moleküler kütlelere sahip küçük ısıya dayanıklı antimikrobiyal peptitlerdir ve durgunluk fazı sırasında ribozomal yolla sentezlenir. Çoğunlukla diğer bakterilerin büyümesini ve (veya) canlılığını etkilerler (de la Fuente-Salcido ve ark. 2013). Bazı çalışmalar, Bt suşlarında sporülasyon ve Cry sentezi sırasında bakteriyosin üretimini bildirmiştir. Şimdiye kadar, Bt morrisoni, kurstaki, kenyae, entomocidus, tolworthi, tochigiensis ve thuringiensis gibi Bt alt türlerinden (vejetatif büyüme döneminde) 18 farklı bakteriosin türü izole edilmiş ve saflaştırılmıştır. Bt bakteriyosinler, geniş veya dar bir bakterisidal veya bakteriyostatik etki gösterebilir (de la Fuente-Salcido ve ark. 2013).
")
Bitki patojeni bakterilerin antagonisti olarak Bt Bazı Bt suşları tarafından üretilen N-acylhomoserine lactone (AHL) parçalayıcı enzim (Aii. A), biber bitkisinin kök sistemindeki yumuşak çürümenin nedensel ajanı olan Erwinia carotovora gibi patojenik bakterilerin virülansını hafifletebilir. Agrobacterium tumefaciens (Bacthuricin F 103; Kamoun ve ark. 2011), Pseudomonas syringae, Pseudomonas gibi farklı bitki patojenik bakterilere karşı Bt türevi bakteriyosinlerin antibakteriyel aktiviteleri bildirilmiştir (Hong ve diğerleri 2015; Jeong ve diğerleri 2016).
")
Bitki patojeni bakterilerin antagonisti olarak Bt Ayrıca, diğer bakteriyel (Citrobacter çiftçisi ve Streptomyces avermectinius) ve mantar (Paecilomyces variotii, Trichoderma parareesei TPJS-1 ve Trichoderma viride TVJ-S-1) antagonistleri ile karışımlarda Bt (vejetatif hücreler) varlığı, kontrol etkinliklerini önemli ölçüde artırmıştır. Bora vd. (2015), Bt, T. parareesei ve T. viride kombinasyonunun, diğer tedaviler ve kontrole kıyasla R. solanacearum'a karşı maksimum antagonistik etki (% 91. 47) gösterdiğini bildirmiştir. Başka bir çalışmada, domates köklerinin Bt CR-371 ve S. avermectinius ile tedavisi, bakteriyel solgunluk hastalıklarını (R. solanacearum'un neden olduğu) ve kök nematod hastalıklarını (Elsharkawy ve ark. 2015) bastırmıştır.

İnsan ve hayvan patojen bakterilerinin kontrolünde ajan olarak Bt Bazı Bt bakteriyosinleri, farklı insan veya hayvan patojenik bakterilere karşı geleneksel antibiyotik tedavisi için mükemmel alternatifler olarak kullanılmak üzere yüksek potansiyele sahiptir. Ayrıca, enterotoksijenik bakterilerin büyümesini engellemek ve gıdaların raf ömrünü uzatmak için gıda ambalajında biyolojik olarak parçalanabilen doğal ve güvenli gıda koruyucuları olarak da kullanılabilirler. Bt bakteriyosinlerinin nisin ile kombinasyonu, antibakteriyel aktivitelerini geliştirebilir.

İnsan ve hayvan patojen bakterilerinin kontrolünde ajan olarak Bt Örneğin, bir Bt fengisin benzeri lipopeptit, E. coli ve Staphylococcus epidermidis'e karşı antibakteriyel aktivite göstermiştir (Roy ve diğerleri, 2013). Meksika Bt alt türleri morrisoni, kurstaki, kenyae, entomocidus ve tolworthi tarafından üretilen bazı bakteriyosin benzeri bileşikler, insanda kusturucu, ishal ve ölümcül sendromların ajanları olan Bacillus cereus ve Vibrio cholerae'ye karşı yüksek düzeyde aktivite göstermiştir. insanlar (Barboza-Corona ve diğerleri 2007). Bacthuricin F 103, Thuricin S ve Thurincin H, Listeria monocytogenes ve B. cereus'a karşı yüksek antibakteriyel aktivite gösterir ve Thuricin 7, Bacillus weihenstephanensis'in neden olduğu çiğ süt ve süt ürünlerinin bozulmasını önlemiştir (Cherif ve ark. 2001).
, gelişmiş fizikokimyasal")
Metal nanopartikülle rin biyosentezi için bir kaynak olarak Bt Metal nanopartiküller (NP'ler), gelişmiş fizikokimyasal özellikleri ve farklı endüstrilerdeki geniş uygulamaları nedeniyle dikkatleri üzerine çekmiştir. Bakteriler, funguslar, bitki özleri ve diğer biyolojik bazlı ürünler gibi çeşitli biyolojik sistemler, farklı metal NP'lerin sentezi için kullanılmıştır (Juibari ve diğerleri 2011, 2015; Okafor ve diğerleri 2013). Mikroplar kullanılarak sentezlenen NP'ler, temiz, toksik olmayan ve çevre dostu olma gibi önemli avantajlar gösterir ve aynı zamanda ortam sıcaklığında ve basıncında da sentelenebilir. NP'leri üretmek için birkaç bakteri ve fungus türü kullanılmıştır (Das ve diğerleri 2014 a, b; Nayak ve diğerleri 2016).

Metal nanopartikülle rin biyosentezi için bir kaynak olarak Bt Son zamanlarda yapılan bazı araştırmalar, Bt suşlarının gümüş (Banu ve diğerleri 2014; Jain ve diğerleri 2010; Nayak ve diğerleri 2016) ve kobalt (Marimuthu ve diğerleri 2013) gibi metal NP'ler üretme yeteneklerini kanıtlamıştır. Jain vd. (2010) ilk kez Bt'nin spor-kristal karışımını kullanarak yüksek verimli gümüş NP yeşil sentezini bildirmiştir. Ortalama parçacık boyutu, karışık (kübik ve altıgen) yapı ile yaklaşık 15 nm idi. Ag. NP'lerin, E. coli, P. aeroginosa ve S. aureus dahil olmak üzere farklı çoklu ilaca dirençli insan patojenik bakterilere karşı oldukça toksik olduğu bulunmuştur.

Metal nanopartikülle rin biyosentezi için bir kaynak olarak Bt Bazı bakterilerin, metal iyonlarının nanopartiküllere indirgenmesini içeren indirgeyici enzimler içerdiği daha önce bildirilmiştir. Bu nedenle, bazı Bt suşlarının NP biyosentezi için indirgeyici enzimler içerdiği sonucuna varılabilir. Marimuthu vd. (2013), bir Bt suşu kullanarak kobalt nanopartikül biyosentezini (Co-NP'ler) rapor etmiş ve Co-NP'lerin sıtma vektörüne, Anopheles subpictus'a ve dang vektörü Aedes aegypti'ye (Diptera: Culicidae) karşı yüksek larvisidal aktiviteye sahip olduğunu doğrulamıştır.

Metal nanopartikülle rin biyosentezi için bir kaynak olarak Bt Bazı bakterilerin, metal iyonlarının nanopartiküllere indirgenmesini içeren indirgeyici enzimler içerdiği daha önce bildirilmiştir. Bu nedenle, bazı Bt suşlarının NP biyosentezi için indirgeyici enzimler içerdiği sonucuna varılabilir. Marimuthu vd. (2013), bir Bt suşu kullanarak kobalt nanopartikül biyosentezini (Co-NP'ler) rapor etmiş ve Co-NP'lerin sıtma vektörüne, Anopheles subpictus'a ve dang vektörü Aedes aegypti'ye (Diptera: Culicidae) karşı yüksek larvisidal aktiviteye sahip olduğunu doğrulamıştır. Mikroorganizmalar ve bitki özleri kullanılarak nanopartiküllerin biyosentezi, mekanik veya kimyasal sentezinkinden daha maliyetli olduğundan, NP'lerin biyosentezi için uygun maliyetli proses tasarlama çabaları devam edecektir.

Ağır metallerin bioremediasyo nu için ajan olarak Bt ve kirlilikler Ağır metaller, böcek ilaçları, herbisitler ve petrol türevleri, günümüzde çevre ve insan sağlığı endişelerinin ana kaynağıdır. Bu bileşikler besin zincirinde kolaylıkla birikebilir ve sonuç olarak daha yüksek trofik seviyelerde tehlikelere neden olabilir. Bazı Bt suşları, bazı toksik kirleticileri verimli bir şekilde ayrıştırır. Bu türler toksik ağır metalleri biriktirebilir, bozabilir veya mineralize edebilir.

Daha önce, arsenik, kadmiyum, kurşun, bakır, nikel, çinko, krom, cıva ve uranyumun Bt bazlı biyoremediasyonu rapor edilmiştir (Tablo 4). Ağır metallerin bioremediasyo nu için ajan olarak Bt ve kirlilikler Ayrıca, bazı Bt türleri, fenantren, imidakloprid, fipronil, klorpirifos sihalotrin, fenoksibenzoik asit, trifeniltin (bir organotin herbisit), difeniltin ve monofeniltin gibi kalıcı pestisitleri ve herbisitleri parçalayabilir. Ayrıca, petrol kirliliğinin (dizel yakıtı ve ham petrol), polisiklik aromatik hidrokarbonların (floranten ve piren), boyaların (metilen mavisi ve asit kırmızısı), organik atıkların (damıtma atığı), malakit yeşili ve melanoidinler ve ayrıca plastikleştirici malzemelerin (dimetil ftalat) Bt temelli verimli bir şekilde bozunmasına dair raporlar literatürde mevcuttur (Tablo 4). Bu bulgular Bt suşlarının gelecekte biyoremediasyon projelerinde önemli bir yer bulacağını göstermektedir. Bununla birlikte, biyoremediasyon amaçlı Bt bazlı ticari bir ürün yoktur ve bu nedenle, bu ürünlerin ticarileştirilmesine yol açmak için daha fazla araştırma ve geliştirme projesi yürütmek gerekir.

Bt nin antikanser özellikleri Cry toksinleri öncelikle Bt tarafından üretilen insektisidal toksinler ailesi olarak bilinir. Bununla birlikte, parasporinler (PS'ler) olarak adlandırılan Cry 31 A, Cry 45 A, Cry 46 A, Cry 63 A ve Cry 64 A gibi bazı Bt Cry proteinleri, herhangi bir insektisidal ve hemolitik aktivite göstermezler, ancak yine de proteazlarla sindirildiklerinde normal hücrelere zarar vermeksizin insan kanser hücrelerine karşı güçlü sitotoksik aktiviteye sahiptir. Parasporin Sınıflandırma ve İsimlendirme Komitesi, amino asit sekans homolojilerine göre altı alt sınıfta (PS 1, PS 2, PS 3, PS 4, PS 5 ve PS 6) gruplandırılan 19 farklı parasporini kaydetmiştir. Parasporinlerin antikanser aktiviteleri İnsan rahim ağzı kanseri hücreleri (He. La (Brasseur ve Parasporinlerin antikanser aktiviteleri, insan rahim ağzı kanseri hücreleri, murin lenfoma L 5178 YR hücre hattı, insan lösemi T hücreleri, insan uterus endometriyum adenokarsinom hücre hatları, miyeloid lösemi hücreleri (HL 60) ve karaciğer (hepatosit) kanseri gibi farklı kanser hücrelerine karşı doğrulanmıştır.

Bt nin antikanser özellikleri Son zamanlarda, kanser insidansındaki önemli artış ve mevcut tedavi yöntemlerinin sınırlamaları, bilim insanlarını yeni etkili terapötik ajanlar bulmak için yoğun araştırma projeleri yapmaya itmiştir. Parasporinler, hedeflenen antikanser tedavisi için potansiyel adaylar olarak bilindiğinden, muhtemelen reseptör aracılığı yoluyla olan etki tarzlarının karakterizasyonu önemlidir. Bilinen parasporinler, çeşitli kanser hücre dizilerine karşı farklı bir etki modu sergiler. PS-1, apoptoz sinyallerini aktive ederek ve Ca 2 + konsantrasyonunu artırarak kanser hücresi ölümünü indükler. He. La hücre hattındaki beklin-1, PS-1'in reseptörü olarak işlev görür.

Bt nin antikanser özellikleri PS-2, gözenek oluşturan bir toksindir ve kanser hücresi plazma membranını hedef alarak bir sitolizin görevi görür. Bu parasporinin yapısı ve işlevi böcek öldürücü Cry proteinlerine benzer ve bu nedenle oligomerizasyonu ve kanser hücrelerinde gözenek oluşumu için glikosilfosfatidilinositol-bağlantılı proteinler. PS-3 ve PS-6, böcek öldürücü Cry toksinlerine benzer üç alanlı yapıya sahiptir. Ayrıca, kanser hücresi plazma zarını etkileyen gözenek oluşturucu bir toksin görevi de görebilirler. PS 4, plazma zarına spesifik olmayan bağlanarak ve hedef hücrede oligomerik kompleksler oluşturarak kanser hücrelerini öldürür.

Bt nin antikanser özellikleri Şu anda, piyasada antikanser biyo ürünleri olarak Bt bazlı farmasötikler yoktur, ancak etki mekanizmalarının belirlenmesi ile, Bt-parasporinlerin gelecekte antikanser ilaçları olarak kullanımı mümkün olabilir.
- Slides: 55