Metabolismo de lipdios MAIOR FONTE DE RESERVA ENERGTICA

Metabolismo de lipídios MAIOR FONTE DE RESERVA ENERGÉTICA

Degradação de Gorduras - Triacilgliceróis As gorduras estão concentradas nos adipócitos Triacilgliceróis Ácidos Graxos + Glicerol LIPASES Os ácidos graxos e o glicerol são liberados pela corrente sanguínea e são absorvidos por outras células (principalmente hepatócitos) O glicerol é convertido em gliceraldeído-3 P, que é processado pela glicólise ou neoglicogênese. Os ácidos graxos são oxidados a Acetil-Co. A que pode seguir o Ciclo de Krebs ou ser convertido em corpos cetônicos.
 Triacilglicerol Lipases Ácido Graxo Glicerol Acetil-Co. A Gliceraldeído 3 P")
Degradação de Triacilgliceróis (Gorduras) Triacilglicerol Lipases Ácido Graxo Glicerol Acetil-Co. A Gliceraldeído 3 P Ácido cítrico Acetoacetato Glicólise Gliconeogênese Ciclo de Krebs Corpos Cetônicos

Conversão de glicerol em gliceraldeído-3 P Glicerol + ATP 1 Glicerol 3 -fosfato + ADP Glicerol 3 -fosfato + NAD+ 2 Dihidroxiacetona fosfato + NADH + H Dihidroxiacetona fosfato 3 Gliceraldeído 3 fosfato 1 Glicerol quinase 2 Glicerol 3 fosfato desidrogenase 3 Triose fosfato isomerase +
 são")
Metabolismo dos Ácidos Graxos Na célula hepática, os ácidos graxos (de cadeia longa) são ativados para formar Acil-Co. A (membrana mitocondrial externa) Em seguida, são transportados para dentro da mitocôndria por um carreador – CARNITINA Na mitocôndria ocorre a oxidação até Acetil-Co. A

Ácidos Graxos de Cadeia Longa Ácidos Graxos de Cadeia Curta (difundem-se pela membrana da mitocôndria) Ativação Transporte Beta-oxidação

Ativação do ácido graxo Na membrana mitocondrial externa. Ácido graxo + ATP + Co-A Acil-Co. A + AMP + PPi Acil-Co. A sintetase

Ligação do Ácido Graxo à Carnitina na membrana mitocondrial interna Acil-Co. A + Carnitina Acil-Carnitina + Co. A Carnitina Acil transferase I e II Carnitina é um derivado da Lisina. Encontrada na carne vermelha Pessoas com baixos níveis de carnitina muitas vezes têm depósito de gordura nos músculos, são irritáveis e fracas. Acil-Co. A Carnitina Acil- Carnitina Membrana da Mitocôndria Acil-Co. A

A carnitina aciltransferase I é inibida pelo malonil-Co. A. O malonil-Co. A é um intermediário da biossíntese de ácidos graxos. A concentração de malonil-Co. A é alta quando a biossíntese de ácidos graxos está acontecendo no citosol. A biossíntese de ácidos graxos só ocorre quando houver excesso de glicídeos, de energia e escassez de ácidos graxos. Assim, quando a síntese de ácidos graxos estiver ocorrendo, a degradação é inibida.

Oxidação de Ácidos Graxos – Matriz Mitocondrial Processo conhecido como b-oxidação ou Ciclo de Lynen Na b -oxidação, os ácidos graxos originam acetil-Co. A. O processo envolve 4 etapas: • Desidrogenação • Hidratação • Oxidação • Tiólise

Beta-oxidação FADH 2 • Desidrogenação • Hidratação NADH + H • Oxidação • Tiólise

A cada ciclo de beta-oxidação, origina-se UMA molécula de Acetil Co. A e tem-se a redução de UMA molécula de NAD+ e UMA molécula de FAD+. Notar que a beta-oxidação de um ácido graxo de n Carbonos, originará 1/2 n moléculas de Acetil Co. A, (1/2 n – 1) moléculas de NADH e (1/2 n – 1) moléculas de FADH 2 Por exemplo, a beta-oxidação de um ácido graxo de 16 Carbonos irá gerar 8 moléculas de Acetil Co. A e 7 moléculas de NADH e 7 moléculas de FADH 2.

Degradação de Ácidos Graxos Triacilglicerol Lipases Ácido Graxo Glicerol Acetil-Co. A Gliceraldeído 3 P Ácido cítrico Acetoacetato Glicólise Gliconeogênese Ciclo de Krebs CICLO DE KREBS Corpos Cetônicos

O acetil-Co. A formado pela Beta-oxidação dos ácidos graxos só entra para o Ciclo de Krebs se a degradação de lipídeos e carboidratos estiver equilibrada. A entrada do acetil-Co. Ano ciclo de Krebs depende da disponibilidade de oxalacetato. A concentração de oxalacetato diminui muito quando não há glicídeos disponíveis. O oxalacetato é normalmente formado a partir do piruvato (produto final da glicólise em aerobiose), por ação da piruvato carboxilase.

No jejum prolongado, o oxalacetato entra para a gliconeogênese e não estará disponível para condensar com o acetil-Co. A. Nestas condições, o acetil-Co. A é desviado para a formação de corpos cetônicos. O que são Corpos Cetônicos?

Corpos Cetônicos são derivados do Acetil-Co. A O fígado é o principal local de síntese de corpos cetônicos. A produção de corpos cetônicos é um mecanismo importante de sobrevivência. A córtex adrenal e o músculo cardíaco utilizam corpos cetônicos (acetoacetato) preferencialmente como combustíveis celulares. No jejum prolongado, o cérebro se adapta à utilização de corpos cetônicos como combustível celular.

Pulmões Acetona não é utilizada pelo organismo e é expelida pelos pulmões Uma indicação que uma pessoa está produzindo corpos cetônicos é a presença de acetona em sua respiração. Acetoacetato e beta-hidroxibutirato podem ser convertidos novamente a acetil-Co. A.

Corpos Cetônicos são produzidos em pequenas quantidades por pessoas sadias. A concentração no sangue de mamíferos normais é de cerca de 1 mg/d. L. A perda urinária no homem é de menos que 1 mg/24 horas. Em algumas condições como jejum ou diabetes, corpos cetônicos atingem altos níveis, acarretando cetonemia e cetonúria. O quadro geral é denominado cetose. O ácido acetoacético e hidroxi-butírico são ácidos moderadamente fortes e precisam ser neutralizados. A excreção urinária desses ácidos provoca acidez da urina. Os rins produzem amônia para neutralizar esta acidez, resultando em diminuição da reserva alcalina - “cetoacidose”.

REGULAÇÃO DO METABOLISMO DE TRIACILGLICEROIS DEGRADAÇÃO DE GORDURAS Com baixa ingestão calórica ou glicemia baixa, ocorre liberação de Glucagon Durante a atividade física ocorre liberação de Epinefrina AMBOS HORMÔNIOS ESTIMULAM A DEGRADAÇÃO DE TRIACILGLICERÓIS Glucagon – TECIDO ADIPOSO Epinefrina – MÚSCULO Glucagon e Epinefrina promovem a degradação de triacilgliceróis pela cascata do c. AMP, fosforilando Lipases.
 Adenilato ciclase a ((ativa ) (inativa) ATP c AMP")
HORMONAL Hormônio (epinefrina ou glucagon) Adenilato ciclase a ((ativa ) (inativa) ATP c AMP + PP Ativação Proteina kinase (inativa ) Proteina kinase (ativa) ATP ADP Lipase ( (inativa) Lipase (P) (ativa) Aumenta degradação de triacilgliceróis

Insulina – é liberada quando a glicemia é ELEVADA Promove a desfosforilação das Lipases. Portanto: INIBE A DEGRADAÇÃO DE TRIACILGLICERÓIS
 Hormônio (epinefrina ou glucagon) Adenilato ciclase (active) (inativa) ATP Proteínas Fosfatases")
HORMONAL Hormônio (Insulina) Hormônio (epinefrina ou glucagon) Adenilato ciclase (active) (inativa) ATP Proteínas Fosfatases c AMP + PP Ativação Proteina kinase (inativa ) Proteina kinase (ativa) ATP ADP Lipase ( (inativa) Lipase (P) (ativa) P INIBE A DEGRADAÇÃO DE TRIACILGLICEROIS

LEMBRETE 1 Lipogênese Quando ingerimos carboidratos, estes são transformados em glicose, e esta, entra na corrente sanguínea. Quando a concentração de glicose depositada no sangue ultrapassa o seu limite máximo, seu excesso é removido pelo fígado, e este, o armazena em seu interior sob a forma de glicogênio, logo ingerindo glicose, consequentemente aumentamos a concentração de glicogênio dentro do fígado. Por sua vez, quando em excesso, o glicogênio é quebrado pelo fígado tendo seu excedente eliminado no sangue e, consequentemente, a concentração de ácidos graxos na corrente sanguínea será aumentada.

LEMBRETE 2 O excesso de ácidos graxos no sangue é removido pela pele, e esta, o armazenará dentro de células dos adipócitos. Esse armazenamento ocorrerá sob a forma de gordura. Lipólise A lipólise é exatamente o processo contrário da lipogênese. Quando o sangue está com concentração de glicose abaixo do normal, ele recebe glicose do fígado resultante da quebra do glicogênio. O fígado, por sua vez, para manter seu nível de glicogênio estável, retira ácidos graxos do sangue, transformando-os em glicogênio.

LEMBRETE 3 Quando o sangue, que teve os ácidos graxos removidos pelo fígado, chega até a pele, esta, quebra a gordura armazenada em seus adipócitos e a introduz no sangue sob a forma de ácidos graxos. Os fatores que influenciam a lipólise e a lipogênese são: ingestão calórica, gasto energético, hormonal, fatores psicológicos, sóciofamiliares e hereditário.
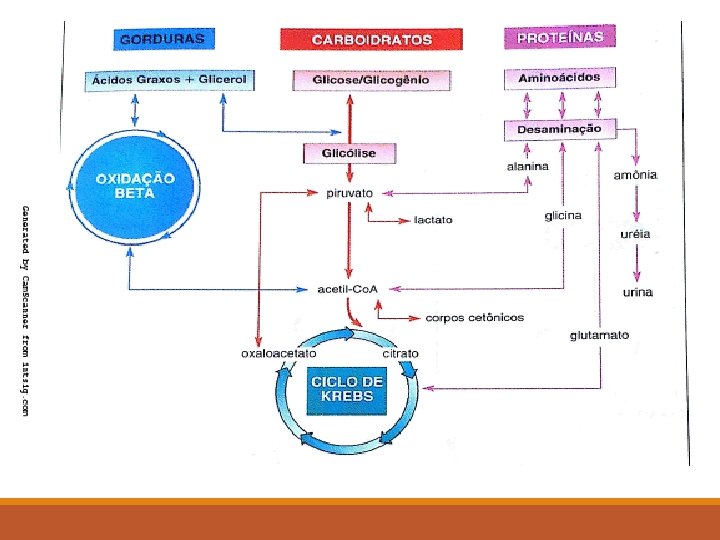

LEMBRETE 4 A maior parte da reserva energética do organismo encontra-se armazenada sob a forma de triacilglicéridos. Estes podem ser hidrolisados por lipase à glicerol e ácidos graxos. O glicerol é oxidado à diidroxiacetona fosfato. A diidroxiacetona fosfato faz parte na seqüência da glicólise. Esse composto pode ser convertido em glicogênio no fígado e tecidos musculares ou em ácido pirúvico, o qual entra no Ciclo de Krebs. Já os ácidos graxos têm como “destino” a β-oxidação.

Resumo de Beta-oxidação dos ácidos graxos É o processo pelo qual o ácido graxo é convertido em acetil-Co. A para a entrada no ciclo de Krebs, acontece dentro da mitocôndria. Nesse processo a βoxidação remove dois átomos de carbono da cadeia de ácido graxo. Os ácidos graxos livres podem passar para dentro da célula por difusão simples pela membrana plasmática, porém não podem entrar livremente para o interior das mitocôndrias. A entrada dos ácidos graxos no interior das mitocôndrias requer primeiro a transformação dos ácidos graxos em acil-Co. A. A membrana da mitocôndria é impermeável á acil-Co. A. Para entrarem na mitocôndria estes reagem com um aminoácido "especial", a carnitina, liberando a coenzima A. A carnitina esterificada é transportada para dentro da mitocôndria por um transportador específico; a carnitina livre volta então para o citoplasma através do transportador. Neste processo não existe transporte de Co. A para dentro da mitocôndria: as reservas citoplasmática e mitocondrial de Co. A não se misturam.

A β-oxidação dos ácidos graxos consiste num ciclo de três reações sucessivas, idênticas à parte final do ciclo de Krebs Por ação da enzima tiolase, libera-se acetil-Co. A, e um acil-Co. A com menos dois carbonos que o acil-Co. A original. A repetição do ciclo permite a degradação total de um ácido graxo de cadeia par em acetil-Co. A, que pode entrar no ciclo de Krebs, onde é completamente oxidado a CO 2; sendo assim é impossível utilizar acetil-Co. A para produzir oxaloacetato.
- Slides: 29