Metabolismo de Glcidos Metabolismo de Glcidos Reacciones con

Metabolismo de Glúcidos

Metabolismo de Glúcidos Reacciones con las que los seres vivos adquieren, producen y utilizan energía para sus diferentes funciones. Tiene cuatro funciones específicas: 1. Obtenergía química de la degradación de los nutrientes. 2. Convertir las moléculas nutrientes en precursores. 3. Sintetizar las macromoléculas biológicas necesarias para la célula. 4. Sintetizar o degradar biomoléculas, necesarias para ciertas funciones celulares.
 conjunto de reacciones por las que la célula degrada los")
Rutas metabólicas CATABOLISMO: (degradación) conjunto de reacciones por las que la célula degrada los nutrientes ANABOLISMO: (biosíntesis) reacciones mediante las que la célula sintetiza sus biomoléculas Las moléculas reaccionantes, intermediarios y productos, se denominan metabolitos o, también intermediarios metabólicos.

Catabolismo de biomoléculas La mayor parte de las rutas catabólicas aerobias de glúcidos, lípidos y proteínas convergen en unos pocos productos finales. Puede considerarse tres etapas: 1. Degradación de las macromoléculas en sus unidades constitutivas. 2. Degradación de esas unidades en moléculas más simples: Pyr y Ac. Co. A 3. Oxidación total de esas unidades en el ciclo del ácido cítrico (Krebs)

Las vías catabólicas aerobias convergen todas en el ciclo de Krebs, que es uno de los puntos claves del metabolismo celular.

Metabolismo de Carbohidratos • • • Glucolisis Fermentación Transformación del piruvato en Acetil-Co. A Ciclo de Krebs (ácidos tricarboxílicos o ácido cítrico) Transporte electrónico y fosforilación oxidativa

Glucólisis Consiste en una secuencia de 10 reacciones enzimáticas que catalizan la transformación de una molécula de glucosa a dos de piruvato, con la producción de dos moles de ATP y dos de NADH por mol de glucosa Prepara la glucosa y otros carbohidratos para su posterior degradación oxidativa. La vía glucolítica implica la transformación de la glucosa en piruvato; el piruvato formado puede sufrir posteriormente distintas degradaciones, dependiendo de las condiciones y del organismo

En condiciones aerobias, el piruvato se transforma en Acetil-Co. A, que se oxida aun más a través del ciclo del ácido cítrico, y posteriormente a través de la fosforilacion oxidativa, generando CO 2 y agua. En condiciones anaerobias tiene lugar la fermentación; la transformación del piruvato hasta moléculas con un grado medio de oxidación. Dos de las fermentaciones más importantes son la homoláctica donde el piruvato es reducido a lactato, y la alcohólica, en levaduras, por la que se reduce hasta etanol y CO 2.

La glucolisis convierte la molécula de glucosa en dos de piruvato, en un proceso que utiliza la energía libre liberada para sintetizar ATP a partir de ADP y fosfato inorgánico (Pi).
. Fase preparatoria en que la glucosa es fosforilada")
• FASE I. (Reacciones 15). Fase preparatoria en que la glucosa es fosforilada y fragmentada, dando lugar a dos moléculas de gliceraldehido-3 fosfato. Este proceso consume 2 ATPs. • FASE II (Reacciones 6 -10). Las dos moléculas anteriormente formadas se convierten a dos moléculas de piruvato, con la producción de 4 ATPs y 2 NADH.

Reacciones de la Glucólisis 1. Consumo del primer ATP Transferencia del grupo fosforilo del ATP a la glucosa para formar glucosa-6 -fosfato (G 6 P) en una reacción catalizada por la hexoquina. La glucosa es fosforilada en el carbono 6
 catalizada")
2. Isomerización Conversión de G 6 P a Fructosa-6 -fosfato (F 6 P) catalizada por la Fosfoglucosa isomerasa. Primero debe abrirse el anillo para que ocurra la isomerización, con posterior ciclación de la fructosa. Para la apertura del anillo se requiere la presencia de un grupo ácido, probablemente el resto de butilamonio de una lisina.

3. Consumo del segundo ATP La fosfofructoquinasa fosforila la F 6 P para formar fructosa 1, 6 -bifosfato (FBP). Esta reacción controla la velocidad de la vía glucolítica. Esta reacción es estimulada alostéricamente por AMP e inhibida alostéricamente por ATP y citrato.

4. Rotura La aldolasa cataliza la rotura de la FBP en dos triosas, el gliceraldehido-3 -fosfato (GAP) y la dihidroxíacetonafosfato (DHAP). Dos moléculas de 3 carbonos

5. Isomerización Sólo uno de los productos de la rotura aldólica, el GAP, continua la vía glucolítica. La interconversión entre éste y la DHAP es catalizada por la triosa fosfato isomerasa. Termina 1 ra fase - 2 ATP

6. Formación del primer intermediario de "alta energía“ La gliceraldehido-3 -fosfato deshidrogenasa cataliza la oxidación y fosforilación del GAP, por Nicotinamida Adenina Dinucleotido (NAD+) y fosfato inorgánico, para producir el 1, 3 -bifosfoglicerato (BFG). Fosfato inorgánico Cada GAP es oxidado y fosforilado por fosfato inorgánico

El NAD+ es el principal agente oxidante de la vía glucolítica, así que el NADH formado durante el proceso debe ser continuamente reoxidado para mantener el suministro de NAD+.

7. Primera producción de ATP Se forma el primer ATP por defosforilación del 1, 3 bisfosfoglicerato, rindiendo además 3 -fosfoglicerato (3 PG) en una reacción catalizada por la fosfoglicerato quinasa (PGK).

8. Isomerización La fosfoglicerato mutasa cataliza la conversión de 3 PG a 2 -fosfoglicerato (2 PG ) Cambia de posición el grupo fosfato

9. Formación del segundo intermediario de "alta energía“ La enolasa cataliza la deshidratación del 2 PG a fosfoenolpiruvato (PEP), formando un complejo activo por la presencia del catión magnesio.

10. Producción del segundo ATP La piruvato quinasa cataliza el acoplamiento de la energía libre de la hidrólisis del PEP a la síntesis de ATP para formar piruvato. Se forman 2 moleculas netas de ATP por 1 molécula de glucosa

El rendimiento de la glucolisis es de dos ATPs formados por molécula de glucosa y la reacción global sería: Glucosa + 2 NAD+ + 2 ADP + 2 Piruvato + 2 NADH + 2 ATP + 2 H 2 O + 4 H+ Desde un punto de vista global podemos decir que la glucólisis se inhibe cuando hay mucho ATP. Los puntos clave en la regulación de la glucólisis son las tres enzimas que catalizan pasos irreversibles: la hexoquinasa, la fosfofructokinasa y la piruvato kinasa.

Otros azúcares en la glicólisis Además de la glucosa procedente de la degradación de almidón y glucógeno, hay otras hexosas de importancia, como la fructosa, que procede de la hidrólisis del azúcar de mesa y también de la fruta, la galactosa, que procede de la hidrólisis del azúcar de leche (lactosa), y la manosa, obtenida a partir de la digestión de polisacáridos y glucoproteínas. La fructosa es fosforilada en el músculo y convertida directamente a F 6 P, siguiendo después la vía glucolítica gracias a la acción de la hexoquinasa. No obstante, en el hígado la fructosa sigue una ruta más compleja cuyo resultado final es la producción de dos unidades de GAP que se incorpora a la ruta.

La galactosa se transforma en glucosa-6 -fosfato, aunque este proceso parece simple las enzimas de la glucolisis no son capaces de reconocer la configuración de la galactosa, lo que hace que el proceso sea catalizado por 5 enzimas. La manosa es fosforilada para rendir manosa-6 -fosfato y a continuación se produce una isomerización hasta fructosa-6 -fosfato
 + 2 H+ + 2 e- NADH (reducido) + H+ El")
Fermentación NAD+ (oxidado) + 2 H+ + 2 e- NADH (reducido) + H+ El NAD+ (limitado en la célula) consumido en la glucólisis debe ser reciclado. En presencia de O 2, el NADH pasa a la mitocondria para oxidado. En condición anaerobia, el NAD+ se recupera por reducción del piruvato. La fermentación recupera el NAD+, dos ejemplos son la fermentación homoláctica y la alcohólica. Destinos del piruvato Condiciones anaeróbicas 2 Etanol + 2 CO 2 En levaduras 2 Piruvato 2 Acetil Co. A TCA 4 CO 2 + 4 H 2 O Condiciones anaeróbicas 2 Lactato Músculo contrayéndose vigorosamente
, la demanda de ATP es elevada")
Fermentación homoláctica En el músculo, (en ejercicio intenso), la demanda de ATP es elevada y se consume el oxígeno, la lactato deshidrogenasa (LDH) cataliza la oxidación del NADH por el piruvato para dar lactato. La reacción global de fermentación homoláctica de glucosa: Glucosa + 2 ADP + 2 Pi 2 lactato + 2 ATP + 2 H+

Fermentación alcohólica En levadura, el NAD+ se regenera en condiciones anaeróbicas mediante la conversión de piruvato a etanol y dióxido de carbono. El etanol es el componente activo de vinos y licores, y el CO 2 es el responsable del aumento de volumen del pan. El etanol se produce a través de las siguientes reacciones
 es un tiol cuyos")
Estructura de la Coenzima A La coenzima A (Co. A) es un tiol cuyos tioésteres actúan como reactivos bioquímicos de transferencia del grupo acilo. El acetil-Co. A transfiere un grupo acetilo a un nucleófilo, actuando el coenzima A como grupo saliente.

El flavín adenín dinucleótido al reducirse capta dos protones y dos electrones, pudiendo intervenir como poder reductor en el metabolismo. El FAD oxida los alcanos a alquenos, mientras que el NAD+ oxida los alcoholes a aldehídos o cetonas. La oxidación de un alcano (como el succinato) a un alqueno (como el fumarato) es lo suficientemente exergónica como para reducir el FAD a FADH 2, pero no para reducir el NAD+ a NADH.

Conversión del piruvato en Acetil Co-A Los grupos acetilo entran en el ciclo en forma de acetil-Co. A. Los grupos acetilo son el producto común de la degradación de carbohidratos, ácidos grasos y aminoácidos y entran al ciclo en forma de Acetil Co-A. La hidrólisis del enlace tioéster del acetil-Co. A libera 31, 5 k. J/mol y es, por lo tanto, un enlace rico en energía. El acetil-Co. A se forma por descarboxilación oxidativa del piruvato, por la acción del complejo enzimático piruvato deshidrogenasa.

La piruvato dehidrogenasa se inhibe cuando las proporciones de ATP/ADP y NADH/NAD+ son altas; y cuando la disponibilidad de combustible para el ciclo, en foma de Acetil-Co. A o ácidos grasos, es alta. Se activa cuando las demandas energéticas crecen y por tanto el flujo de Acetil-Co. A aumenta.

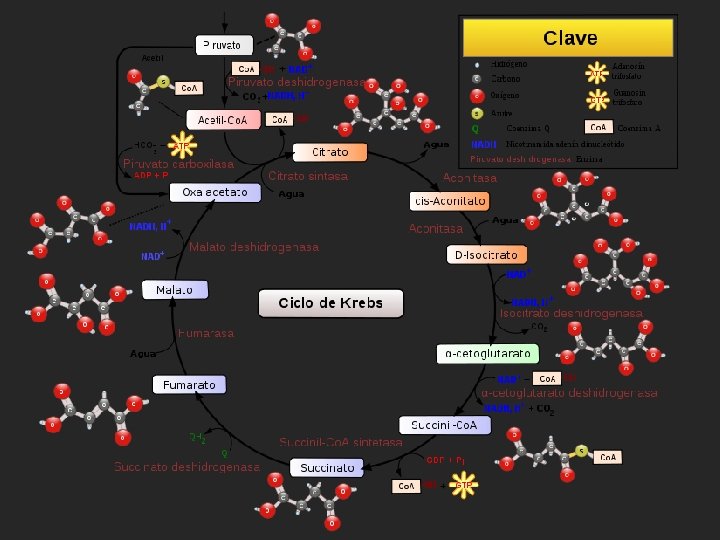
Ciclo de Krebs Es la vía de oxidación de la mayor parte de carbohidratos, ácidos grasos y aminoácidos. Las ocho enzimas del ciclo catalizan una serie de reacciones que oxidan un grupo acetilo a dos moléculas de dióxido de carbono, con la generación de tres moléculas de NADH, una de FADH 2 y una de GTP

Reacciones del ciclo de Krebs 1. La citrato-sintasa cataliza la condensación entre acetil-Co. A y oxalacetato para rendir citrato, que da nombre al ciclo.

2. La aconitasa convierte el citrato en isocitrato que es más fácilmente oxidable, mediante una deshidratación que produce cis-aconitato unido al enzima, seguida de una hidratación. Así, el grupo hidroxilo del citrato es transferido a un átomo de carbono adyacente.

3. La isocitrato deshidrogenasa oxida el isocitrato a αoxalosuccinato, con la reducción acoplada de NAD+ a NADH. Posteriormente, el oxalosuccinato es descarboxilado, rindiendo α-cetoglutarato. Esta es la primera etapa en la que la oxidación se acopla a la producción de NADH, y también la primera en la que se genera dióxido de carbono.

4. El complejo enzimático α-cetoglutarato deshidrogenasa descarboxila oxidativamente el α-cetoglutarato a succinil-Co. A. Esta reacción conlleva la reducción de una segunda molécula de NAD+ a NADH y la generación de una segunda molécula de dióxido de carbono. Hasta aquí ya se han producido dos moléculas de dióxido de carbono, por lo que se ha completado la oxidación neta del grupo acetilo. Hay que resaltar que no son los átomos del grupo acetilo entrante los que han sido oxidados

5. La succinil-Co. A sintetasa convierte el succinil-Co. A en succinato. La energía libre de la reacción se conserva aquí por la formación de GTP, a partir de GDP y Pi. Guanosina trifosfato (GTP) Guanosina difosfato (GDP) Guanosina monofosfato (GMP)

6. La succinato deshidrogenasa la cuál cataliza la oxidación del enlace sencillo situado en el centro de la molécula de succinato a un doble enlace trans, dando lugar a fumarato con la reducción simultánea de FAD a FADH 2.

7. La fumarasa cataliza después la hidratación del doble enlace del fumarato para rendir malato.

8. Finalmente, la enzima malato deshidrogenasa regenera el oxalacetato, oxidando el grupo alcohol secundario del malato a la correspondiente cetona, con la reducción de una tercera molécula de NAD+ a NADH.

La energía de las oxidaciones se conserva con eficiencia Con un ciclo tiene lugar: • 2 átomos de carbono entran al ciclo y se combinan con el oxalacetato que viene del ciclo anterior. • 2 átomos de carbono salen del ciclo en forma de CO 2 en los procesos de oxidación del isocitrato y el α-cetoglutarato • Finalmente se regenera la molécula de oxalacetato. para el ciclo siguiente. • Los átomos de carbono que aparecen como CO 2, no son los mismos que entran en forma de grupo acetilo. • Se requieren varios ciclos para que los átomos de carbono del grupo acetilo salgan en forma de CO 2

La oxidación completa de los grupos acetilo sigue entonces la siguiente estequiometría 3 NAD+ + FAD + GDP + acetil-Co. A + Pi 3 NADH + FADH 2 + GTP + Co. A + 2 CO 2 La oxidación de un acetilo (2 CO 2) por cada vuelta del ciclo, genera: 3 NADH, 1 FADH 2, 1 GTP (o ATP)

Transporte electrónico La glucosa se oxida a CO 2 mediante las reacciones de glucolisis y ciclo de Krebs. La oxidación completa de la glucosa se escribe como indica la siguiente ecuación: Glucosa + 6 O 2 6 CO 2 + 6 H 2 O Separando en dos semirreacciones, podemos expresar en la primera la oxidación de los átomos de C y en la segunda la reducción del oxígeno molecular: C 6 H 12 O 6 + 6 H 2 O 6 CO 2 + 24 H+ + 24 e- 6 O 2 + 24 H+ + 24 e- 12 H 2 O

Los 12 pares de electrones involucrados en la oxidación de la glucosa no pasan directamente al oxígeno, sino que se transfieren a los coenzimas NAD+ y FAD, formándose un total de 10 NADH y 2 FADH 2 Glucólisis (2 ATP + 2 NADH) 2 + (2 x 3) = 8 ATP 6 Descarboxilación oxidativa del piruvato = 6 ATP 2 GTP Ciclo de Krebs (2 ATP + 6 NADH + 2 FADH 2 ) 2 + (6 x 3) + (2 x 2) = 24 ATP 38 Total: 8 + 6 + 24 = 38 ATP. Por cada molecula de Piruvato-Acetil-Co. A: 3 NADH, 1 FADH 2, 1 GTP (ATP) Por dos moleculas: 6 NADH, 2 FADH 2, 2 GTP (o ATP)

La obtención de ATP a partir de la oxidación de NADH y FADH 2 se realiza mediante la fosforilación oxidativa: 1 NADH= 3 ATP 1 FADH 2 = 2 ATP 1 GTP= ATP Los electrones involucrados pasan a la cadena de transporte electrónico donde participan (por la reoxidación mitocondrial del NADH y FADH 2) en un proceso de oxidación -reducción. En este proceso, los protones (H+) son expulsados de la mitocondria, y el gradiente de p. H resultante impulsa la síntesis de ATP, a partir de ADP y Pi, a través de la fosforilación oxidativa.

La reoxidación de cada NADH da lugar a la síntesis de 3 ATP, y la de un FADH 2 a 2 ATP. El total por molécula de glucosa oxidada es pues de 38 ATP, 30 proceden de los 10 NADH, 4 de los 2 FADH 2, además en la glucolisis se producen 2 ATP por mol de glucosa y en el ciclo de Krebs 2 GTP (= 2 ATP) por cada 2 de piruvato que entra en el ciclo. El primer paso es la entrada de los electrones en la cadena respiratoria. La mayoría de los electrones provienen de la acción de dehidrogenasas que recogen los electrones de los distintos procesos catabólicos y los canalizan hacia los aceptores universales de electrones (NAD+, NADP+, FAD).

Entonces los electrones son transferidos a una serie de transportadores asociados a membrana. Estos transportadores son de naturaleza proteica y tiene grupos prostéticos capaces de aceptar/donar electrones. En la cadena respiratoria intervienen tres tipos de moléculas capaces de transportar electrones. La ubiquinona o coenzima Q (una quinona hidrofóbica), los citocromos (proteinas que tienen como grupos prostéticos grupos hemo con hierro) y las proteínas con agrupaciones sulfo-férricas.
 transporta los electrones del NADH a la ubiquinona.")
• El complejo I, (NADH-ubiquinona-oxidorreductasa) transporta los electrones del NADH a la ubiquinona. • El complejo II, (succinato-dehidrogenasa), única enzima del ciclo de Krebs unida a membrana, que pasa los electrones del FADH 2 a la ubiquinona. • El complejo III (citocromo bc 1 ó ubiquinona-citocromo coxidorreductasa), acopla la transferencia de electrones desde la ubiquinona al citocromo c. • El complejo IV, (citocromo-oxidasa), es la última etapa de la cadena de transporte electrónico de la respiración y conduce los electrones desde el citocromo c hasta el último aceptor de los electrones, el oxígeno que se reduce a agua
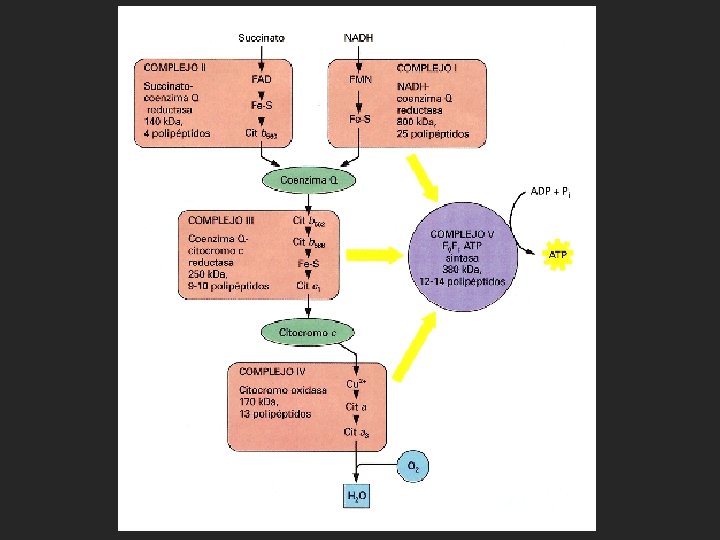

• Fosforilación oxidativa: La síntesis de ATP a partir de ADP y Pi en las mitocondrias está catalizada por la ATP sintasa (complejo V), y está impulsada mediante el proceso de transporte electrónico anterior. La energía liberada durante el transporte debe conservarse en una forma que pueda ser usada por la ATP-sintasa. Esto se conoce como acoplamiento de energía o transducción de energía.

La teoría propone que los transportadores de electrones además de transportar electrones bombean protones desde la matriz mitocondrial al espacio intermembrana en contra del gradiente, para ser llevado a cabo este proceso endergónico es acoplado a la energía producida por el transporte de electrones a favor de gradiente, de modo que se crea un gradiente electroquímico de protones a través de la membrana mitocondrial interna. El potencial electroquímico de este gradiente es aprovechado por la ATP sintasa para sintetizar ATP. La ATP sintasa transporta los protones a la matriz mitocondrial a favor de gradiente y acopla este proceso exergónico a al síntesis de ATP

De esta forma, el transporte electrónico provoca que los complejos I, III y IV transporten protones a través de la membrana mitocondrial interna desde la matriz (una región de baja concentración de protones y potencial eléctrico negativo), al espacio intermembranal (una región de elevada concentración de protones y potencial eléctrico positivo).

Respiraciones no aerobias En la respiración aerobia el aceptor final de los electrones es el oxígeno que se reduce a agua. Pero hay organismos que son capaces de respirar sin oxígeno llevando los electrones hasta otros aceptores con el mismo objetivo final, obtener mucho ATP. • Hay organismos capaces de respirrar: • Nitrato, generando nitrógeno (bacterias denitrificantes) • Sulfato, generando sulfuro (bacterias sulforeductoras) • CO 2, generando metano (bacterias metanogénicas)
- Slides: 54