METABOLISMO DE CARBOHIDRATOS Monogstricos Rumiantes LICZA PADILLA Ph

METABOLISMO DE CARBOHIDRATOS Monogástricos- Rumiantes LICZA PADILLA Ph. D

FUENTES DE CARBOHIDRATO

QUE SON LOS CARBOHIDRATOS? • Son polihidroxialdehidos y polihidroxicetonas o compuestos que por hidrólisis se producen. • También son llamados glúcidos o azucares. • Sirven como FUENTE DE ENERGIA para todas las actividades vitales celulares. • En general son moléculas de almacenamiento de energía en la mayoría de los seres vivos en forma de almidón en las plantas o de glucosa en los animales. • Actualmente se acepta denominarlos como glúcidos. • Función estructural (pared celular, exoesqueleto de los artrópodos).

CLASIFICACIÓN DE CARBOHIDRATOS

Nomenclatura Monosacáridos: están formados por una sola molécula. Según el grupo funcional • Aldosas: (poseen como grupo funcional un aldehido) • Cetosas: (Poseen como grupo funcional una cetona) Según el número de carbonos • Triosas: poseen 3 carbonos • Tetrosas: poseen 4 carbonos • Pentosas: poseen 5 carbonos • Hexosas: poseen seis carbonos

Combinando el grupo funcional y número de carbonos • Aldotriosa: posee un grupo funcional aldehído y tres carbonos. Ej. D-Gliceraldehido. • Aldopentosa: posee un grupo funcional aldehído y cinco carbonos. Ej. D-Ribosa. • Aldohexosa: poseen un grupo funcional aldehído y seis carbonos. Ej. D-Glucosa. • Cetoriosa: posee un grupo funcional cetona y tres carbonos. Ej. Dihidroxicetona. • Cetohexosa: posee un grupo funcional cetona y seis carbonos. Ej. Fructosa.

ENTRE LOS PRINCIPALES MONOSACARIDOS • Glucosa: es el principal producto de la digestión de almidón en monogastricos, se presenta en sangre, linfa, liquido cerebroespinal. • Fructuosa: se encuentra libre en plantas verdes, semen y sangre de los fetos. • Galactosa: no existe libre y es un producto de las fermentaciones.

DISACARIDOS • Son azúcares compuestos por dos residuos de monosacáridos. • Unidos por un enlace glicosídico. • Con pérdida de una molécula de agua al realizarse dicha unión. • Solubles en agua, dulces cristalizables.

LA MALTOSA se obtiene por hidrólisis del almidón y del glucógeno, ambos polisacáridos utilizados como reserva de azúcares. (Se utiliza para la elaboración de cerveza). * Disacárido formado por dos glucosas. α- D- glucopiranosil [1 -4] Dglucopiranosa.

La isomaltosa • Se obtiene por hidrólisis de la amilopectina, componente del almidón y del glucógeno. α- D- glucopiranosil [1 -6] Dglucopiranosa.

La sacarosa es el azúcar que consumimos habitualmente. Se obtiene principalmente de la caña de azúcar y de la remolacha azucarera. El producto se obtiene en forma cristalizada procesando esos vegetales. α-D-glucopiranosil-(1 -2)--D-fructofuranósido.

LACTOSA La lactosa se encuentra en la leche de los mamíferos, siendo ésta su única fuente natural. (Se considera de vital importancia durante la lactancia tanto como fuente de energia como de desarrollo inmumologico de las crías). β-D- galactopiranosil [1 – 4] Dglucopiranosa.

POLISACÁRIDOS DE RESERVA Y ESTRUCTURALES Polisacárido de Estructura reserva Caracteristicas Observaciones Almidón Mezcla en proporcion variable de AMILOSA LINEAL y AMILOPECTINA RAMIFICADA Fuentes: se encuentra en las semillas de las plantas y tuberculos como papas y legumbres. Se deposita en las células de estos vegetales formando gránulos. Formado por la unión de muchas moléculas: α-D-glucopiranosa, mediante enlaces α(1 -4) y α(1 -6) De importancia especial las semillas de los cereales. Los granos de almidón están compuestos por amilosa en su parte interior y amilopectina en sus capas externas. Ambos compuestos son hidrolizados por enzimas amilasas.

Polisacárido de reserva Estructura Características Glucógeno Es ramificado con estructura semejante a la de amilopectina, pero con ramificaciones mas próximas, cada 4 a 8 moléculas de glucosa. Es el polímero de reserva en los animales, se almacena en todos los tejidos, pero especialmente en músculo e hígado. Ramificaciones secundarias, terciarias y cuaternarias numerosas. Observaciones La glucosa producida durante la fotosíntesis en las plantas verdes e ingerida por los animales con los alimentos, debe almacenarse inmediatamente.

GLUCÓGENO

Polisacarido reserva Estructura Caracteristica Observaciones Inulina Es un polisacarido lineal formado por monomeros de Fructosa con enlaces glucosidicos. β-(2 -1) Forma parte de Se almacena en la fibra de la tuberculos, raices dieta. de dalias, alcachofas y diente de leon

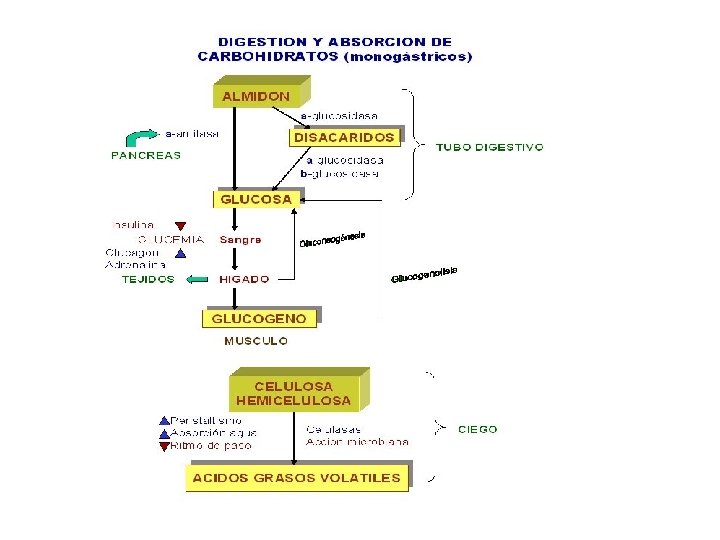
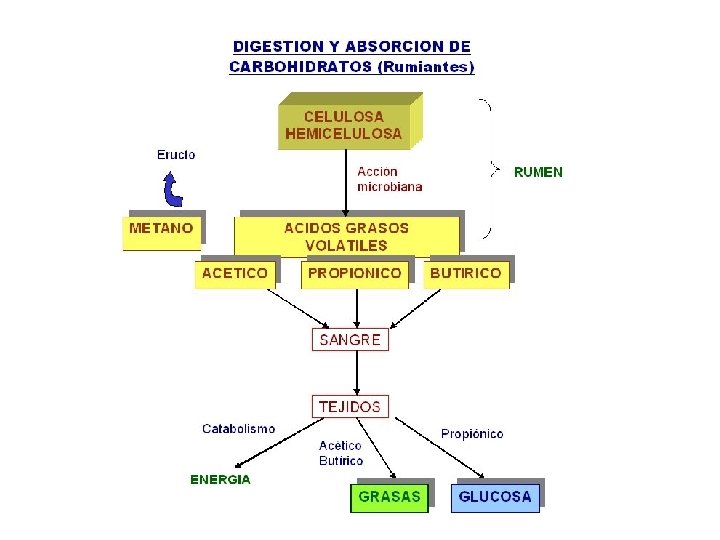
Polisacarido Estructural Celulosa Estructura Caracteristicas Observaciones Es un homopolímero lineal. No forma helices, su cadena es mas menos rectilinea. Esta constituido por mas de 10, 000 moleculas de Glucosa unidas por enlaces β- (1 -4) Es el compuesto orgánico mas abundante en la naturaleza. Los organismos superiores no poseen enzimas capaces de catalizar la hidrólisis de las uniones β-(1 -4), por tal razon no se pueden utilizar como alimento. - En el intestino de los rumiantes y otros herbívoros existen microorganismos capaces de hidrolizar estos enlaces , haciendo disponible la celulosa como fuente calórica importante para tales animales. - El producto final de la digestión de celulosa, es una mezcla de ácidos grasos volátiles y gases como metano y C 02. Forma las paredes celulares de los vegetales.

Polisacáridos de origen vegetal • La pectina, que se encuentra en la pared celular de los tejidos vegetales. Abunda en la manzana, pera, ciruela, membrillo y cascara de naranja. Posee una gran capacidad gelificante que se aprovecha para preparar las mermeladas. • Hemicelulosa: polisacáridos ramificados y lineales con restos de azucares como: xilosa, arabinosa, glucosa, galactosa y ac. propionico. Se encuentra en la pared celular de plantas forrajeras. • El agar-agar, que se extrae de las algas rojas o rodofíceas. Es muy hidrófilo y se utiliza en Microbiología para preparar medios de cultivo. • La goma arábiga, sustancia segregada por las plantas y que sirve para cerrar sus heridas. • Lignina : Es una sustancia que aparece en los tejidos leñosos de los vegetales y que mantiene unidas las fibras de celulosa que los componen, la lignina constituye el 25% de la madera. (Complejo aromático)

Contenido de carbohidratos estructurales en forrajes Carbohidrato Porcentaje Celulosa 20 -40 Hemicelulosa 10 -40 Lignina 5 -10 Pectina 1 -10

Contenido de glicógeno en el hígado de los animales Especie Glicógeno en el hígado (%) Perros 4. 3 Ovejos 3. 82 Vacas (lactantes) 1. 0 Vacas (Secas) 3. 0 Lechones 5. 2 Lechones (recién nacidos) 14. 8

Digestión de Carbohidratos • En los animales, la digestión de los carbohidratos comienza con el contacto del alimento con las enzimas presentes en la saliva. • El almidón contenido en las plantas y el glicógeno contenido en la carne son fragmentados a monosacáridos por acción de la amilasa salival (ptialina) y maltasa. • Esta actividad cesa cuando la materia alimenticia pasa al estómago, donde la actividad enzimática es desactivada por acción del ácido clorhídrico. • En el estómago ocurre la hidrólisis ácida, sin embargo el estómago se vacía rápidamente sin completarla. Por lo tanto solamente una porción de los carbohidratos ingeridos es hidrolizado antes de pasar al intestino delgado.

Digestión de Carbohidratos- Monogástricos • En el intestino delgado la digestión de los carbohidratos se lleva a cabo rápidamente por enzimas fragmentadoras de carbohidratos contenidas en los jugos pancreáticos (amilasa pancreática o amilopsina) y los jugos intestinales. • El almidón y el glicógeno son hidrolizados a glucosa por acción de la amilasa y la maltasa; la lactosa es hidrolizada a glucosa y galactosa por acción de la lactasa y la sacarosa a glucosa y fructosa por acción de la sacarasa. • Los monosacáridos productos de la hidrólisis enzimática de los carbohidratos: glucosa, fructosa y galactosa. Son la principal forma de absorción en animales monogástricos.
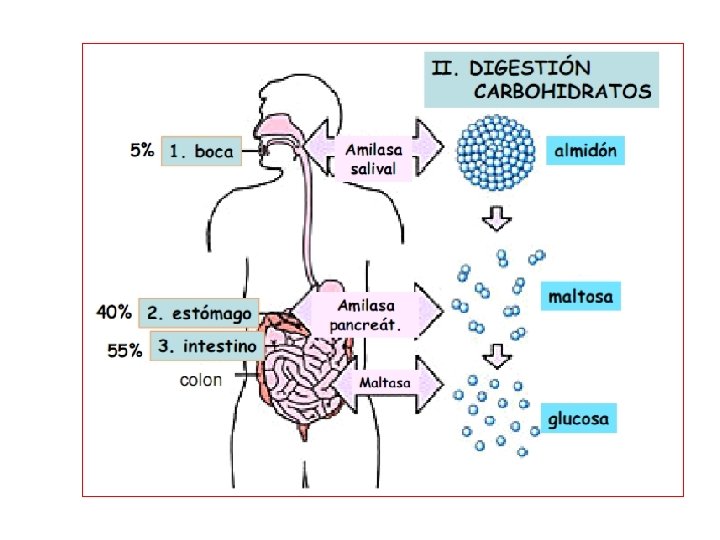

Absorción de los Carbohidratos-Monogástricos • Los monosacáridos son absorbidos casi por completo a través de la mucosa del intestino delgado y se encuentran en la circulación como azúcar libre. • La glucosa y la galactosa son absorbidas rápidamente. • Mientras la fructosa es absorbida aproximadamente a la mitad de la tasa de absorción de la glucosa, siendo una parte convertida a glucosa en el proceso. • Otros monosacáridos son absorbidos lentamente, como resultado, azucares libres aparecen en la circulación portal para su transporte al hígado.

• La absorción en la célula ocurre por un mecanismo de transporte activo sodio dependiente utilizando un co-transportador de glucosa. • La glucosa absorbida sale de la célula mediante el proceso de difusión en presencia del transportador de glucosa: GLUT-2.

El paso de los monosacáridos son conducidos por la vena porta hacia el hígado donde la galactosa y la fructosa son convertidas a glucosa. La glucosa es el único azúcar que circula en la sangre: azúcar de la sangre. La glucosa entonces, puede: (1) pasar al sistema circulatorio general para ser transportada a los tejidos o bien (2) incorporarse al glicogeno para quedar almacenada como reserva a fin de mantener el nivel normal de glucosa en la sangre.

En los tejidos: • La glucosa puede oxidarse a CO 2 y H 2 O. • Convertirse en grasa. • Convertirse en glicógeno muscular y servir como fuente de energía fácilmente disponible para el músculo. Una persona normal posee una cantidad almacenada de glicógeno para suministrar energía por ± 18 h.

Destino de los carbohidratos GLUCOSA GALACTOSA FRUCTOSA - Fuente inmediata de energía - Glucógeno hepático y muscular - Triglicéridos tisulares

Glucosa de la sangre • La concentracion de glucosa en la sangre o Nivel de azucar en la sangre, es vital para el funcionamiento del cuerpo. Condiciones normales, nivel de azucar en la sangre es relativamente constante 80 mg/100 ml de sangre. (5 -6 g de glucosa o una cucharada en todo el cuerpo) Hipoglicemia: Concentración de glucosa inferior a la normal. Los casos extremos se deben a cantidades excesivas de insulina, puede causar inconsciencia, presión sanguínea baja y provocar la muerte. Hiperglicemia: Concentración de glucosa superior a la normal. Casos extremos se alcanza el umbral renal 160 -170 mg/100 ml y el exceso de glucosa es excretado por la orina

Sintesis y descomposición del glicogeno • Es el proceso mas importante que contribuye mantenimiento de un nivel constante de glucosa. • Algunas hormonas regulan el nivel de azucar en la sangre • El pancreas secreta la insulina y su funcion es la de reducir el nivel de azucar en la sangre. Haciendo mas permeables las membranas celulares al paso de la glucosa e incrementando la glicogenesis. al

Glicogénesis: Formación de glicógeno a partir de la glucosa Gliconeogénesis Formación de glicogeno a partir de fuentes diferentes a las de los carbohidratos (aminoácidos, piruvato) Glicogenolisis La adrenalina (epinefrina) activa a la enzima fosforilasa, la cual tiene relación con la degradación del glicógeno a glucosa Condición diabética: Ocurre cuando el pancreas no secreta la suficiente cantidad de insulina o por problemas en los receptores celulares. Todas las demas hormonas que afectan al metabolismo del azucar actuan para elevar la concentracion de azucar en la sangre.
 Glucosa sanguínea Glicógeno Adrenalina (glicogenólisis)")
Insulina (glicogenesis) Glucosa sanguínea Glicógeno Adrenalina (glicogenólisis)

*Existe almacenamiento de glucosa disponible en el hígado en forma de glicógeno que puede ser liberada por este órgano para que otros tejidos la puedan utilizar como fuente de energía. *La segunda fuente importante de almacenamiento de glucosa es el glicógeno del músculo esquelético. Sin embargo, el glicógeno del músculo no esta disponible para otros tejidos, debido a que el músculo carece de la enzima glucosa-6 fosfatasa. El glicógeno almacenado en el hígado es considerado como el principal amortiguador "buffer" de los niveles de glucosa de la sangre.

Metabolismos de los carbohidratos absorbidos • Las células del hígado son fácilmente permeables a la glucosa. • Este proceso es facilitado por las proteínas transportadoras de glucosa. Las proteínas que se encuentran en la membrana plasmática, en particular GLUT 2 son las transportadoras en las células del hígado. En el hígado, existen diferentes vías que determinan el destino inmediato de las hexosas. • La glucosa, fructosa y la galactosa forman glucosa fosfatada entonces, • Los fosfatos de glucosa son convertidos y almacenados como glicógeno, catabolizados a CO 2 y agua o a azucares libres retornando a la circulación.

METABOLISMO DE LA GLUCOSA Vía del metabolismo de la glucosa-6 -fosfato La conversión fundamental que requiere la iniciación de la oxidación de la glucosa en la célula es la fosforilación a la forma G-6 -P. Existen por lo menos cinco diferentes vías que la G-6 -P puede seguir: • A. Glucosa libre • B. Glicólisis • C. Glicogenesis • D. Hexosa monofosfato • F. y la vía del glucoranato

A. GLUCOSA LIBRE G-6 -fosfatasa Glucosa -6 -PO 4 Glucosa + Pi • Esta es una reacción irreversible opuesta a la reacción de la glucocinasa (GK). • Estas dos opuestas e independientes reacciones de enzimas catabolizadoras son el sitio del control metabólico de la glucosa ya que el balance de la actividad enzimática regula la dirección de las reacciones.

• Significantes cantidades de G-6 -fosfatasa son encontradas en el hígado y en menor cantidad en los riñones. (no se encuentra en el músculo). • La actividad de la G-6 -fosfatasa es generalmente mayor que la GK por casi las 24 horas del día exceptuando en unas pocas horas después de la ingesta de alimentos. Esto significa que la mayor parte del tiempo el hígado suple glucosa en vez de usar glucosa.

B. Glicólisis Anaerobia/Trayectoria de Embden-Meyerhof Los bioquímicos alemanes Gustav Embden y Otto Meyerhof dilucidaron la secuencia de reacciones mediante las cuales el GLICÓGENO y la GLUCOSA, son degradados en ausencia de oxigeno (condiciones anaerobias) a ÁCIDO PIRÚVICO. Se descubrió que la fermentación alcohólica con levaduras y la contracción muscular en animales ocurría por la misma trayectoria hasta ácido pirúvico. En ausencia de oxigeno Las células musculares (otros tejidos animales) convierten ácido pirúvico a ácido láctico En condiciones semejantes las enzimas de las levaduras transforman el ácido pirúvico a alcohol etílico y dióxido de carbono. Glicólisis Fermentación

La característica sobresaliente trayectoria Embden-Meyerhof es el empleo de la coezima NAD+ como receptora de electrones (agente oxidante).

Glicógeno Glucosa Acido pirúvico

Por lo tanto el ácido láctico es el producto final de la glicólisis y si no hubiera un mecanismo para eliminarlo se acumularía en las células musculares elevando el nivel de acidez en la célula. El aumento de acidez impide la función muscular causando fatiga y agotamientos musculares.

Importancia del ácido pirúvico Las etapas 1 -10 son idénticas para; - La glicólisis - La fermentación.

Factores energéticos de la Glicolisis y de la Fermentación Rendimiento energético neto en forma de enlaces fosfato de alta energía (moles de ATP) que se obtiene del metabolismo anaeróbico de cada mol de glucosa Se consume un mol de ATP en la fosforilación inicial (etapa 1) y otro mol en la fosforilación de fructosa 6 -fosfato (etapa 3). En la etapa 7 cada mol de triosa se convierte en un mol de acido 3 fosfoglicerico; se produce un mol de ATP por mol de triosa, o dos moles de ATP por mol de glucosa. En la etapa 10 se genera un mol de ATP por cada mol de acido pirúvico que se forma a partir del acido fosfoenolpiruvico; nuevamente se producen dos moles de ATP por cada mol de glucosa que participa en la trayectoria.

La suma de todas las etapas revela que por cada mol de glucosa que se degrada, inicialmente se consumen dos moles de ATP y por ultimo se producen cuatro moles de ATP: • La producción neta de ATP es de dos moles por mol de glucosa que se convierte a acido láctico o a etanol (la levadura). • Si se emplea glucógeno como fuente, las etapas i y ii en lugar de ATP (etapa 3) a fin de producir la fructosa 1, 6 -difosfato, y el rendimiento neto seria de tres moles de ATP por mol de glucosa 1 -fosfato (Musculo partiendo del glicógeno).

Almacenamiento de Energía Química El desdoblamiento de del ATP a ADP y fosfato inorgánico proporciona la energía para la contracción muscular. El fosfato de creatina es la forma de almacenamiento de la energía en los músculos de los vertebrados y en el tejido nervioso. El fosfato de arginina sirve para el mismo fin en los músculos de invertebrados. Reciben el nombre de fosfagenos y son fosfatos de alto contenido energético.

La concentración de ATP en el musculo es muy baja, por lo tanto el fosfato de creatina, en presencia de creatina cinasa, reacciona con el ADP produciendo ATP y creatina. Fosfato de creatina + ADP Cinasa Mg 2+ Creatina + ATP Cuando se requiere actividad muscular la reacción sucede hacia la derecha. Cuando hay ATP en abundancia la reacción se efectúa hacia la izquierda.

CICLO DE KREBS

Metabolismo aerobio, Ciclo de Krebs El acido pirúvico primero se descarboxila para formar un compuesto de dos carbonos, el cual entra en una secuencia cíclica de reacciones conocida globalmente como ciclo de Krebs, ciclo del acido tricarboxilico o ciclo del acido cítrico.
 no es intermediario del ciclo de Krebs.")
Etapa 1. El acido pirúvico (3 C) no es intermediario del ciclo de Krebs. Primero se debe descarboxilar y oxidarse enzimaticamente (descarboxilación oxidativa) para producir la acetil Co. A (2 C), intermediario del ciclo Krebs. Se necesitan 6 cofactores: Co. A (B 5) TPP(B 1), acido lipoico, NAD+ (B 3), FAD (B 2) y Mg 2+. 1 er. Esta etapa es irreversible.
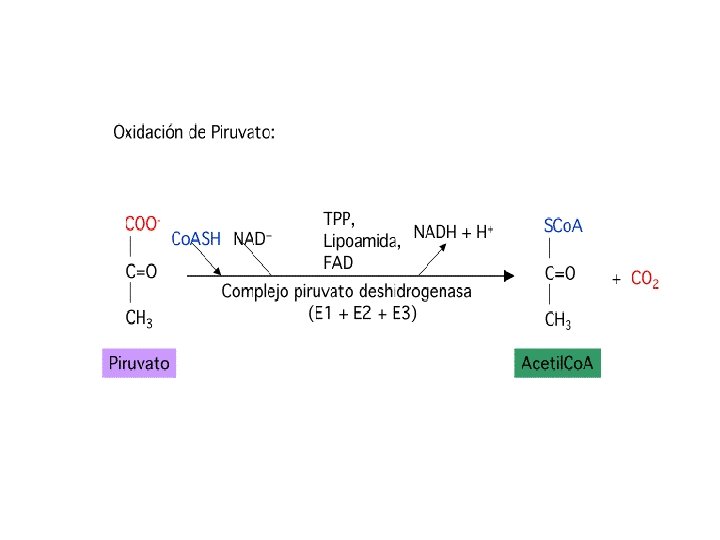

Coenzimas que participan en el complejo PDH tienen relación con vitaminas del complejo B • TPP. La tiamina es la vitamina B 1. Su falta ocasiona el beri (perdida de peso, disfunción neurológica, temblores). • Coenzima A. El ácido pantoténico es la Vitamina B 5. Su falta causa hipertensión. • NAD. El ácido nicotínico(nicotinamida es su derivado) es la vitamina B 3. Su falta ocasiona la pelagra (dermatitis, depresión, diarrea). • FAD. La riboflavinas la vitamina B 2. Su falta ocasiona lesiones bucales, dermatitis.

Etapa 2. Probablemente la acetil Co. A no es un intermediario verdadero del ciclo de Krebs, sino que entra al ciclo al condensarse con el ac. Oxalacetico (4 C) formando ac. Citrico (6 C), al cual se debe el nombre del ciclo. En esta etapa se regenera la COA

Etapa 3. La enzima simple acotinasa cataliza la eliminación y reincorporación sucesiva de una molécula de H 2 O. El resultado neto es la isomeracion del acido cítrico a su isómero menos simétrico, el acido isocitrico.

Etapas 4 y 5. Estas etapas se consideran juntas porque el intermediario, acido oxalsuccinico, no existe en estado libre, sino que unido a la superficie de la enzima. Descarboxilacion del acido isocitrico a acido α-cetoglutarico.

Etapa 6. Esta etapa es prácticamente idéntica a la etapa 1, es catalizada por el complejo acido α-cetoglutarico deshidrogenasa, el cual requiere los mismos 6 cofactores que la ácido piruvico deshidrogenasa. Reacción irreversible del ciclo, lo cual evita que el ciclo opere en dirección inversa.

Etapa 7. Hidrólisis de la succinil Co. A con la formación del trifosfato de guanosina (GTP), a partir de GDP y fosfato inorgánico. Es importante porque el GDP puede reaccionar con el ADP para generar ATP.

Etapa 8. La enzima acido succinico deshidrogenasa cataliza la eliminación de 2 atomos de H del acido succinico formando el acido fumarico. Esta reacción es la única que es catalizada por la coenzima FAD en lugar de la NAD.

Etapa 9. Mediante la adicion de una molecula de H 2 O al doble enlace del acido fumarico catalizada por la enzima fumarasa se forma el acido L -malico.

Etapa 10. La enzima acido málico deshidrogenasa cataliza la deshidrogenación del acido L-málico y se completa una revolución del ciclo.

Cadena respiratoria • Metabolismo aerobio, tiene lugar en presencia de oxigeno molecular. • En el ciclo de Krebs ningún intermediario del ciclo se encuentra unido a grupos fosfato ni tampoco tiene lugar la síntesis directa del ATP a partir de ADP y fosfato inorgánico • El ciclo de Krebs aborda fundamentalmente el destino del esqueleto carbonado del ácido pirúvico describiendo los metabolitos involucrados en su conversión a dioxido de carbono y agua. • Las coenzimas NAD+ y FAD se reducen a NADH y FADH 2. No mecanismo para su regeneración.

Cadena respiratoria o sistema de transporte electrónico Es la secuencia de reacciones mediante la cual se oxidan de nuevo (fundamentalmente mediante oxigeno molecular ) las formas reducidas de las coenzimas.

Balance de ATP en la oxidación de la glucosa Glucosa ATP Fructosa 1 -6 -difosfato -2 2 Piruvatos 1 NADH (3 ATP) 2 ATP 6 4 2 Acetil Co. A 1 NADH (3 ATP) 6 2 Ciclo de Krebs 3 NADH (3 ATP) 1 ATP 1 FADH (2 ATP) 18 2 4 Total 1 NAD utilizado en glicolisis 38 ATP - 2 36 ATP

Control del ciclo de Krebs • Una dieta promedio genera 2000 a 3000 kcal por día (75 g proteina x 4 kcal, 305 g azucar x 4 kcal, 78 g grasa x 9 kcal). • Los cho’s proveen mas del 50% de generación de ATP, y se deben de producir aproximadamente 120 moles de ATP o 65 kg del mismo. • Como el organismo solo tiene 3 a 4 g de nucleotidos (ATP, ADP, AMP) cada molécula debe de ser refosforilada miles de veces al día.
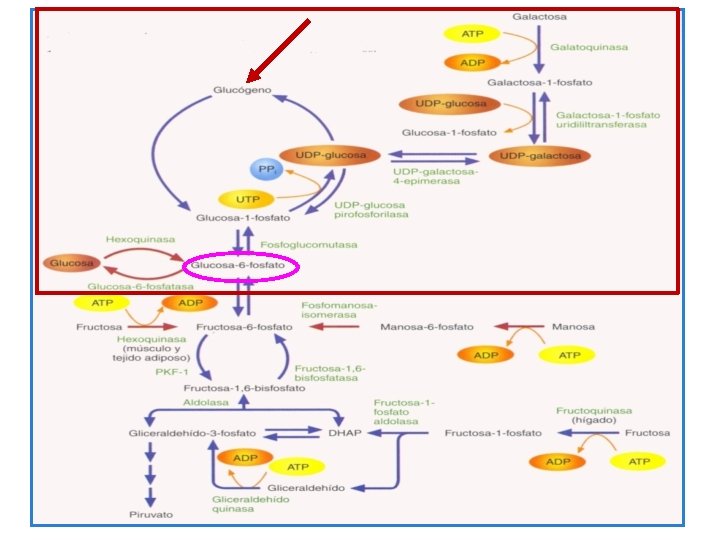

Metabolismo del glicógeno C. Glicogénesis • La reacción inicial requiere la entrada de glucosa en una serie de reacciones metabólicas, las cuales culminan en la síntesis del glicógeno, esto es la fosforilación de la glucosa en el carbono 6. • La glucosa es fosforilada en el hígado mediante ATP, por una reacción enzimática irreversible catalizada por una glucocinasa específica (GK). GK Glucosa +ATP glucosa -6 -P + ADP (I)

• La glucocinasa es una de las cuatro enzimas hexocinasas que se encuentran en todos los tejidos. La GK (HK-IV) específica para glucosa es predominante en el hígado. La no específica (HKI) se encuentra en los glóbulos rojos, cerebro y tejidos nerviosos. • La velocidad de la reacción de fosforilación catalizada por la GK (HK-IV) es controlada por la concentración de glucosa. Su actividad es incrementada mediante la alimentación de glucosa y la insulina y decrece durante el ayuno y la carencia de insulina.

• La inicial unidireccional reacción de fosforilación permite la acumulación de glucosa en las células del hígado, porque los azucares fosforilados no salen o entran libremente a la célula tal y como lo hacen los azúcares libres. • La glucosa-6 -fosfato (G-6 -P) acumulada en la célula experimenta una mutación donde el grupo fosfato es transferido al C-1 de la molécula de glucosa, reacción catalizada por la enzima fosfoglucomutasa (PGM) y envuelve a la glucosa 1 -6 -difosfato como intermediario. Glucosa-6 -P glucosa-1 -P II

• El glicógeno es sintetizado por la G-1 -P mediante una serie de reacciones que envuelven la formación de derivados de la uridina. • Uridina-di-fosfoglucosa (UDP-G) es sintetizada por la transferencia de glucosa de G-1 -P a uridina trifosfato (UTP). Reacción catalizada por la enzima UDP-Gpirofosforilasa (UDP-G-PPasa): UTP + G-1 -P UDP-G + PP (III)
 n (glucosa 1 -4)n+1 +UDP (IV) �")
Glicógeno sintetasa UDP-G + (glucosa 1 -4) n (glucosa 1 -4)n+1 +UDP (IV) � La cadena polisacárida es alargada mediante repetidas transferencias de moléculas de glucosa. Cuando esta alcanza el nivel critico de longitud entre 11 y 16 unidades glucosídicas la enzima para elongación α- glucan glicosil 4: 6 transferasa, forma nuevos enlaces o puntos de ramificación en la cadena de una molécula de glicógeno.

Glicógeno

 ocurre por una")
Glicogenólisis • La fragmentación del glicógeno del hígado a glucosa (glicogenólisis) ocurre por una diferente vía. La clave para la iniciación y regulación de la glicogenólisis es la acción de la Adrenalina (Epinefrina) en el glicógeno del hígado y glicógeno muscular, y la acción del glucagón en el glicógeno del hígado. • El mecanismo de acción del glucagón y la adrenalina se da a través de una serie de reacciones que culminan en la escisión fosforílica del enlace glucosil 1 -4 del glicógeno. • En las células del hígado el glucagón y la adrenalina estimulan la enzima adenil ciclasa para formar 3’-5’ adenosina monofosfato cíclica (c. AMP) desde ATP. c. AMP activa la proteina kinasa quien activa la fosforilasa del hígado (LP).

• La acción del la LP es la escisión del enlace 1 -4 glucosil del glicógeno. • En el hígado la hidrólisis de G-6 -P es catalizada por la enzima glucosa-6 -fosfatasa (G-6 -Pase). Para liberar glucosa libre provocando la hiperglicemia.


• También el glucagón promueve la hiperglicemia mediante la estimulación de la gluconeogénesis. Por lo tanto el glucagón es un importante factor para la hiperglicemia. • Sin embargo en el glicógeno muscular en donde la G-6 fosfatasa se encuentra ausente la fragmentación del glicógeno resulta en la producción y liberación de Piruvato y Lactato en vez de glucosa. • Casi todo el lactato y parte del piruvato es trasportado al hígado donde la glucosa es resintetizada.

CICLO DE CORI

Glicógeno y enfermedades • En enfermedades sistemáticas, cambios en las concentraciones de glicógeno en tejidos u órganos son observados cuando este disminuye. • La reducción de las reservas de glicógeno en el hígado es observada en los casos de diabetes, ayuno y ketosis bovina, o en cualquier condición donde se encuentre deficiencia de carbohidratos.

gluconeogenesis • • Es la producción de azúcares a partir de sustancias diferentes a los carbohidratos (lactato, aminoácidos, propionato y glicerol). Esta vía permite tener una fuente alterna de glucosa, remover el lactato (producidos por los glóbulos rojos y el tejido muscular) de la sangre, remover el glicerol producido por el tejido adiposo. Esta vía metabólica se activa ante la disminución de la glucosa sanguínea, su activación es en ayuno en el cerdo 24 h, hombre 8, y en el pollo 2. En el rumiante es una vía constantemente activa. La gluconeogenesis se encuentra bajo control hormonal (insulina, glucagón, adrenalina)

RUTA DE LAS HEXOSAS MONOFOSFATO O PENTOSAS FOSFATO • Esta vía metabólica ni requiere, ni produce ATP. • Se desarrolla en el citoplasma de las células de tejidos con elevada actividad lipogénica (hígado, tejido adiposo, glándula mamaria, cerebro en desarrollo). • a. Productos principales NADH. (agentes reductores; procesos anabólicos). b. Ribosa 5 – fosfato (componente estructural de nucleótidos y ácidos nucleicos). * Los productos finales pueden contener de 3 a 7 átomos de carbono utilizados posteriormente en la glucolisis (triosas fosfato), en la síntesis de aminoácidos, en la síntesis de ácidos nucleicos, NAD, FAD, Co. A.

RUTA DE LAS PENTOSAS FOSFATO RUTA OXIDATIVA Funciones: a. Activa en células que sintetizan cantidades grandes de lípidos; tejido adiposo, corteza suprarrenal, glándula mamaria, hígado. b. Antioxidante potente (impide la oxidación de otras moléculas). Activa en células con riesgo elevado de daño oxidativo (eritrocitos). Ausente en las células musculares (no sintetizan lípidos). Se produce en dos vias: Oxidativa No oxidativa

RUTA DE LAS PENTOSAS FOSFATO VIA OXIDATIVA .

RUTA DE LAS PENTOSAS FOSFATO RUTA NO OXIDATIVA Se produce la isomerización y la Condensación de varias moléculas de azúcar diferentes. Tres intermediarios de este proceso (resultado) útiles en otras rutas. A. Ribosa – 5 – fosfato. B. Fructosa – 6 – fosfato. C. Gliceraldehido – 3 – fosfato. Pueden degradarse para generar energía o convertirse en moléculas precursoras para los procesos de biosíntesis si, la célula requiere mas moléculas de NADPH, de ribosa o, los productos hacia la glucólisis. .

VIA DEL GLUCORANATO • UDP-Glucoronato formado en esta vía es utilizado en una serie de reacciones importantes en los animales como: formación de bilirrubina y síntesis de mucopolisacáridos. • La L- gulonato puede ser convertida a una pentosa Lxilulosa o vitamina C (ácido ascórbico). • La enzima L- Gulonolactona oxidasa esta ausente en humanos y primates por lo tanto vitamina C debe ser suplementada en sus dietas. Gulonolactona oxidasa

Metabolismo de Carbohidratos en Rumiantes

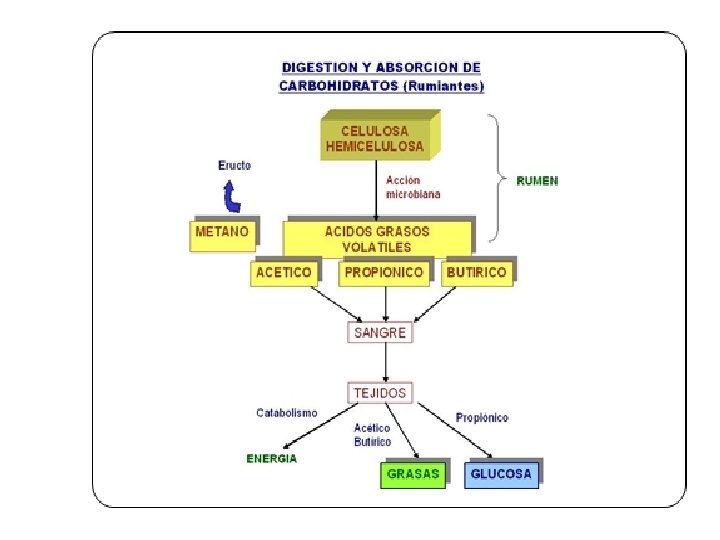
RUMIANTES • La dieta metabólica de los rumiantes es la combinación entre los productos de la fermentación y el alimento no fermentado que escapa a la acción de las bacterias ruminales. • Los rumiantes son eficientes para realizar la gluconeogénesis y su aparato digestivo se ha adaptado a una falta de azúcar y almidón por lo que la capacidad para el manejo de estos carbohidratos es limitada. • Así, los rumiantes absorben la mayoría de su carbono dietario digerido (energía) en forma de ácidos grasos volátiles (AGV).
 • Aumentan")
Función de los carbohidratos fibrosos • Estimulan la rumia (mejoran la fermentación) • Aumentan el flujo de saliva hacia el rumen • Estimulan las contracciones ruminales

Contenido de carbohidratos estructurales en forrajes Carbohidrato Porcentaje Celulosa 20 -40 Hemicelulosa 10 -40 Lignina 5 -10 Pectina 1 -10

Composición de carbohidratos en diferentes ingredientes Ingrediente Mono Di Oligo Poli Almidón Celulosa Hemicelulosa 73 2 3 55 4 10 67 2 8 Pectinas Lignina Cereales Maíz 4 Cebada 1 Trigo Leguminosas Soja Guisantes 5 5 1 7 15 5 -7 2 42 6 15 Vegetativos alfalfa 5 7 14 7 6 6 Gramínea 5 1 25 20 3 8 -12 43 26 3 7 Paja de cebada

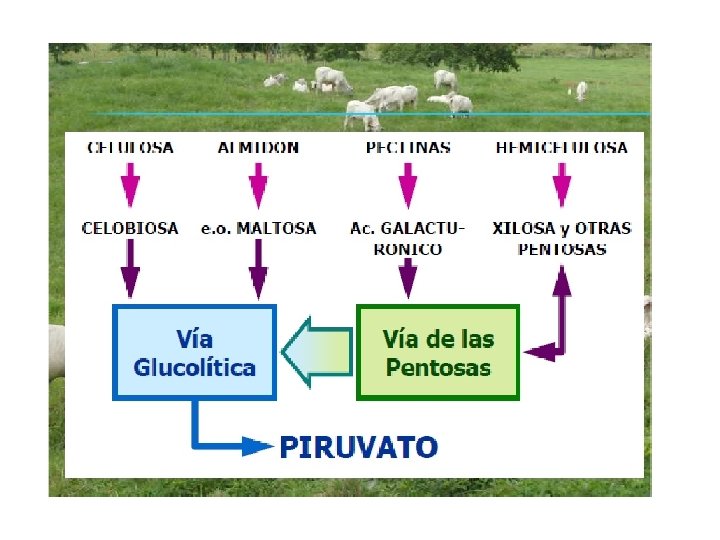
Metabolismo • Todos los carbohidratos independientemente de su clasificación son fermentados en el retículo rumen hasta glucosa y toda glucosa pasa a piruvato. • Por distintas vías metabólicas el piruvato es transformado en acetato, propionato o butirato.
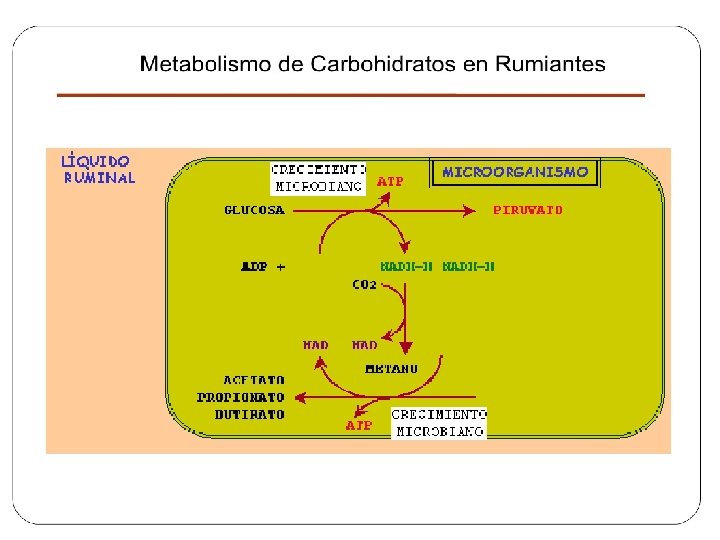

ACIDOS GRASOS VOLATILES PRODUCIDOS EN LA FERMENTACION RUMINAL Nombre Acético Propiónico Butírico Estructura CH 3 -COOH CH 3 -CH 2 -CH 2 -COOH
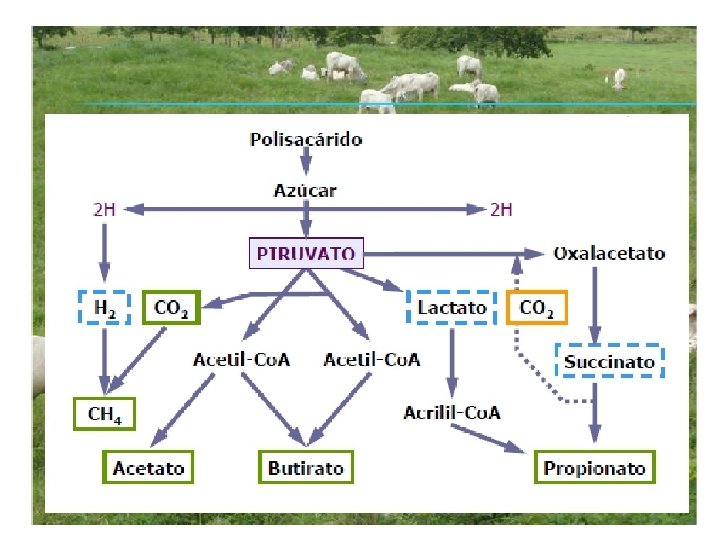

Acido propionico • En los rumiantes la degradación de los carbohidratos en el rumen desprende una gran cantidad de ácido propiónico. • En el hígado el ácido propionico es el único que interviene en la gluconeogenesis. • Hay diferentes vías para llegar al propionato la primera (reductiva directa) donde el piruvato pasa a lactato, el lactato a acrilil Co. A y esta a propionato (sin sufrir decarboxilaciones). • La vía Aleatoria o la vía del oxaloacetato. En este caso se incorpora un carbono al piruvato para formar oxaloacetato que pasa a succinato y de ahí a propionato.

Propionato como fuente de energía El primer paso es la conversión a succinil Co. A.

• El fosfoenol piruvato puede convertirse en fructosadifosfato por inversión de la secuencia de la glicólisis, el cual pasa a fructosa - 6 - fosfato por acción de la difosfructoquinasa y después a glucosa - 6 - fosfato y finalmente a glucosa por la glucosa - 6 - fosfatasa. • La glucosa puede usarse para obtenergía y realizar el siguiente balance:

• Por lo tanto hay una ganancia neta de 17 moles de ATP por cada mol de ácido propiónico. • En la sangre de la circulación sistémica existen pequeñas cantidades de propionato, que pueden proceder de una captura insuficiente por el hígado o de la oxidación de los ácidos grasos de número impar de átomos de carbono. Este propionato puede aprovecharse directamente para la producción de energía.

Ácido butírico como fuente de energía • A su paso a través de las paredes del rumen y del omaso el ácido butírico se convierte en B- hidroxibutirato, puede ser usado como fuente de energía por una serie de tejidos, especialmente el músculo esquelético. • El acetilcoenzima A se metaboliza vía ciclo de los ácidos tricarboxílicos. • Podemos calcular la energía producida a partir del ácido butírico como sigue:

El ácido acético como fuente de energía • El principal producto de la digestión de los carbohidratos en los rumiantes es el ácido acético, que es el único ácido graso volátil que se encuentra en la sangre de la circulación sistémica en cantidades significativas. • Gran número de tejidos lo usan como fuente de energía. • En este caso la primera reacción es la conversión del acetato a acetilcoenzima A en presencia de acetilcoenzima A sintetasa. • El acetilcoenzima A se oxida vía el ciclo de los ácidos tricarboxìlicos produciendo 12 moles de ATP por mol. Puesto que en la reacción inicial mediada por la sintetasa se emplean dos enlaces fosfato de alta energía, la producción neta es de 10 moles de ATP por mol de ácido acético.

Destino de los ácidos grasos volátiles en el rumiante RUMEN EPITELIO RUMINAL VENA PORTA (HIGADO) HIGADO Acetato Propionato Butirato SANGRE Acetato 90% Lactato (15%) Propionato (85%) Gluconeogenesis Glucosa β-OH butirato Butirato Acetoacetato Interconversión β-OH butirato
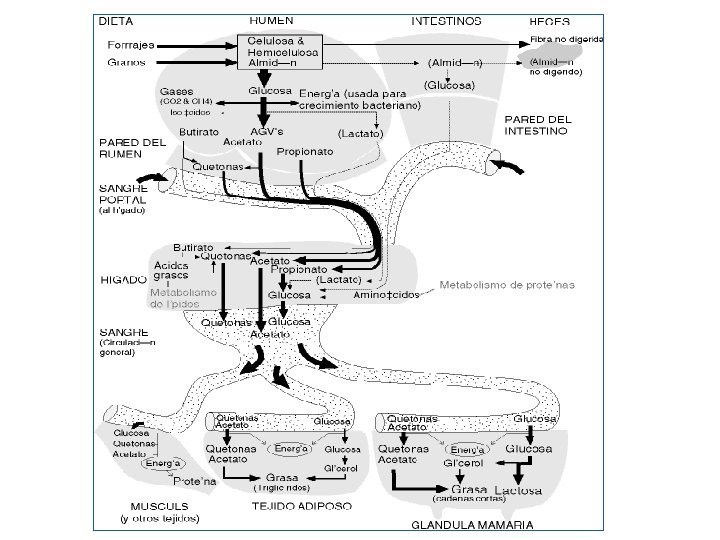

PRODUCCIÓN DE GLUCOSA EN EL HIGADO • Todo el propionato se convierte en glucosa en el hígado. Además, el hígado utiliza los aminoácidos para síntesis de glucosa. • Este es un proceso importante porque normalmente no hay glucosa absorbida del tracto digestivo y todos los azúcares de la leche (aproximadamente 900 g cuando una vaca produce 20 kg de leche) deben ser producidos por el hígado. • Una excepción se produce cuando la vaca se alimenta con grandes cantidades de concentrados ricos en almidón o recibe alguna fuente de almidón resistente a la fermentación ruminal, así este almidón escapa de la fermentación y alcanza el intestino delgado. La glucosa formada mediante la digestión en el intestino es absorbida, y transportada al hígado donde contribuye al suministro de glucosa de la vaca. • El ácido láctico es una fuente alternativa de glucosa para el hígado. El ácido láctico proviene de los ensilajes bien conservados, pero además hay producción de ácido láctico en el rumen hecho que suele ocurrir cuando hay un exceso de almidón en la dieta. Esto no es deseable porque el ambiente del rumen se acidifica, la fermentación de fibra se paraliza y en casos extremos la vaca deja de comer.

SÍNTESIS DE LACTOSA Y GRASA EN EL HIGADO • Durante la lactancia, la glándula mamaria tiene una alta prioridad para la utilización de glucosa. • La glucosa se utiliza principalmente para la formación de lactosa (azúcar en la leche). • La cantidad de lactosa sintetizada en la ubre está estrechamente ligada a la cantidad de leche producida cada día. La concentración de lactosa en la leche es relativamente constante de aproximadamente 4, 5%. Así, la producción de leche en las vacas lecheras está altamente influida por la cantidad de glucosa derivada del propionato producido en el rumen.

SÍNTESIS DE LACTOSA Y GRASA EN EL HIGADO • Los otros dos ácidos, acético y butírico se utilizan para la formación de la grasa de la leche. • También, parte de la glucosa se convierte en glicerol y se utiliza en síntesis de la grasa. La glándula mamaria sintetiza ácidos grasos saturados de cadena corta que contienen de 4 a 16 átomos de carbono. Casi la mitad de grasa de leche es sintetizada en la glándula mamaria. La otra mitad que es rica en ácidos grasos nosaturados que contienen de 16 a 22 átomos de carbono (ácidos grasos de cadena larga) provienen de los lípidos de la dieta.

EFECTO DE LA DIETA SOBRE LA FERMENTACION RUMINAL Y EL RENDIMIENTO DE LECHE • La fuente de carbohidratos del alimento influye sobre la cantidad y la relación de AGV producidos en el rumen. • La población microbiana del rumen genera aproximadamente 65% ácido acético, 20% ácido propionico y 15% ácido butirico cuando la ración contiene una alta proporción de forrajes. • Proporción de AVG con dietas basadas en concentrados: AA: 45%, AP: 40% AB: 15%. • Los carbohidratos no-fibrosos presentes en muchos concentrados promueven la producción de ácido propionico mientras los carbohidratos fibrosos que se encuentran principalmente en forrajes estimulen la producción de ácido acético en el rumen. Además, los carbohidratos no-fibrosos rinden más AGV (es decir más energía) porque son fermentados mas rápidamente y más completamente.
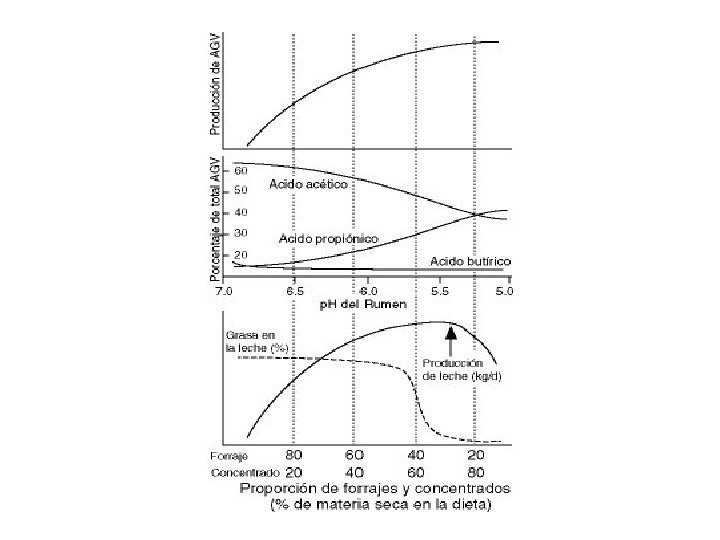

• Un exceso en el concentrado en la ración lleva a vacas gordas. La alimentación prolongada de esta ración puede tener un efecto negativo para la salud de la vaca, que tiene más propensión a padecer hígado graso, cetosis y dificultades en el parto. Por otro lado, cantidades insuficientes de concentrado en la ración limitan la ingestión de energía y la producción de leche. • Un cambio en la proporción de forraje y concentrado en una dieta provoca un cambio importante en las características de los carbohidratos que y tiene un efecto profundo en la cantidad y porcentaje de cada AGV producido en el rumen. En definitiva los AGV tienen un efecto importante sobre: - La producción de leche - El porcentaje de grasa en la leche - La eficiencia de convertir alimentos a leche - El valor relativo de una ración para la producción de leche en lugar de engorde.

Principales problemas que se presentan por la ingesta de carbohidratos • El aumento rápido de los gases puede generar timpanismo consistente en la formación de gran cantidad de espuma estable que llega a bloquear el cardias e impedir el eructo. El origen más común de este trastorno se suele dar de forma crónica, cuando los animales se alimentan con una ración rica en concentrado. • Se puede evitar incluyendo más fibra en la ración y en casos muy graves realizando una punción del rumen. • En otras ocasiones el origen del timpanismo radica en el consumo de pastos de leguminosas aunque en este caso el agente espumante no son los carbohidratos sino la proteína.

• La paraqueratosis del rumen consiste en una queratinización de la mucosa del rumen donde se producen también inflamación y ulceraciones. Por estas penetran bacterias y toxinas que vía sanguínea van al hígado creando abscesos o pueden llegar por el torrente sanguíneo a lugares como las pezuñas y provocar laminitas y cojeras. • La acidosis láctica se produce por una ingestión aguda de concentrados. Aparece de repente mucho ácido láctico generado por la fermentación de carbohidratos fácilmente asimilables con lo que el p. H desciende bruscamente afectando a la microbiología del rumen. Así además de las consecuencias sobre la digestión y fermentación del alimento que conlleva se produce un paso de ácido láctico a sangre y la acidosis se hace sistémica.

Metabolismo anormal de carbohidratos • - Cetosis= Hipoglucemia Acetonuria Exhalación dulce • Diabetes= Hiperglucemia Glucosuria Balance negativo de N

¡GRACIAS POR SU ATENCION!
- Slides: 114