Major histocompatibility Complex T Cell Receptors Janya Nakarin

Major histocompatibility Complex & T Cell Receptors Janya Nakarin Department of Microbiology Faculty of Science Prince of Songkla University janya. n@psu. ac. th

������� . 1��������� MHC ���������� 2. ���������� MHC molecules 3. Antigen processing ��� presentation 4. MHC restriction 5. T cell receptor (TCR) ��� TCR diversity 6. CD 3 complex ��� accessory molecules �� T lymphocyte

A. History 1. In transplantation studies, MHC gene products were identified as responsible for graft rejection 2. In studies on responses to antigens, MHC gene products were found to control immune responses, called the immune response (Ir) genes 3. It was determined that antigenspecific T cells recognize portions of protein antigens that are bound noncovalently to MHC gene products

a. Helper T cells recognize peptide bound to class II MHC gene products b. Cytolytic T cells recognize peptide bound to class I MHC gene products 4. The complete three-dimensional structure for both class I and class II MHC molecules has been
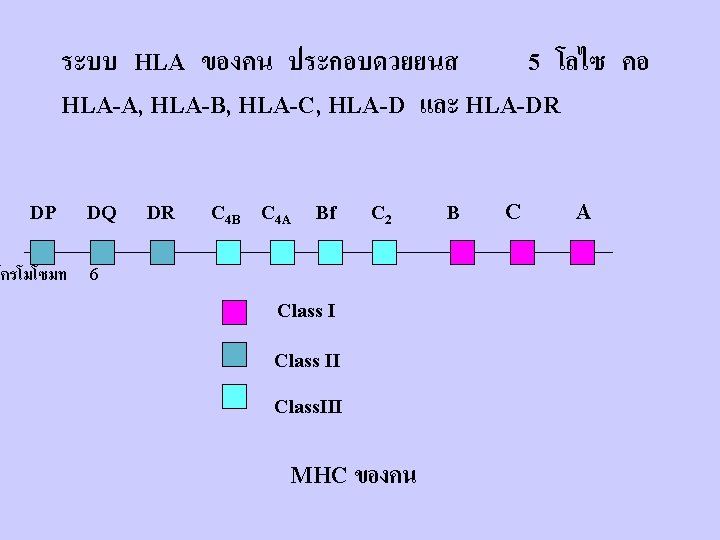

Map of the human MHC

The polymorphic residues of Cl I &Cl II molecules: Cl I : located in the peptide-binding clefts and the a - helices around the clefts(a 1 & a 2). Cl II: show varying degrees of polymorphism in the a chain and usually much more in the b- chain (b 1).

Number of Class I Alleles = 818 Number of Class I I Alleles = 571

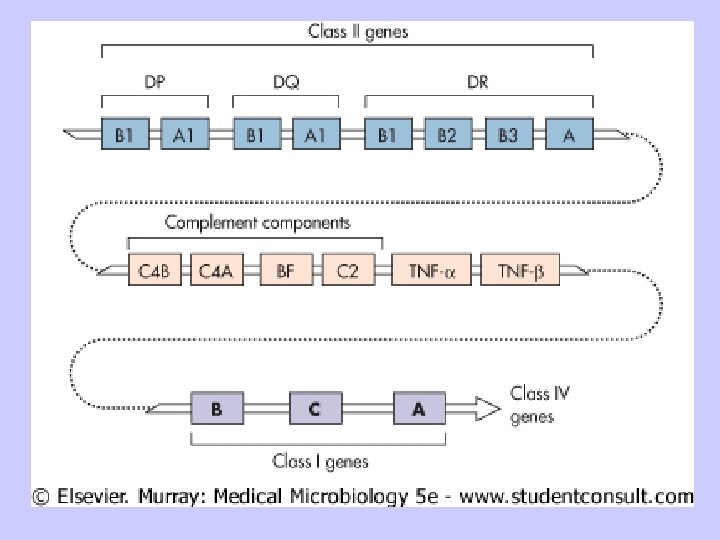
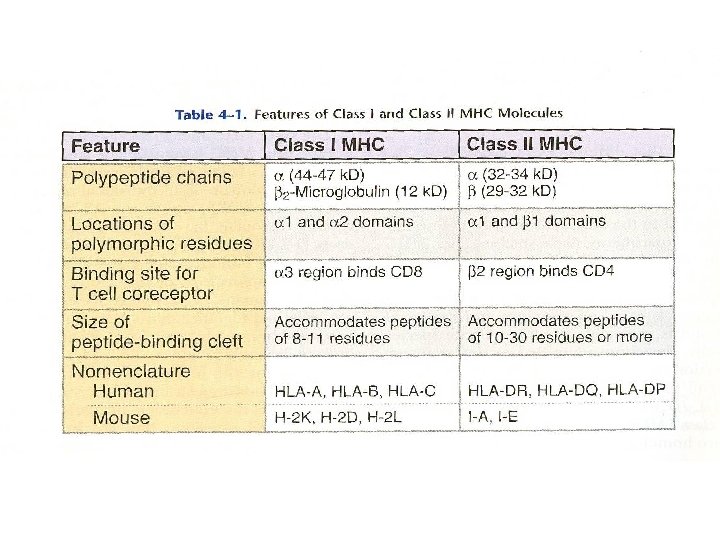
B. Structure of MHC Molecules 1. Class I MHC a. Class I MHC molecules contain two separate polypeptide chains (1) MHC-encoded alpha (or heavy) chain; 43 k. Da (2) non-MHC-encoded beta chain (beta 2 microglobulin) 12 k. Da
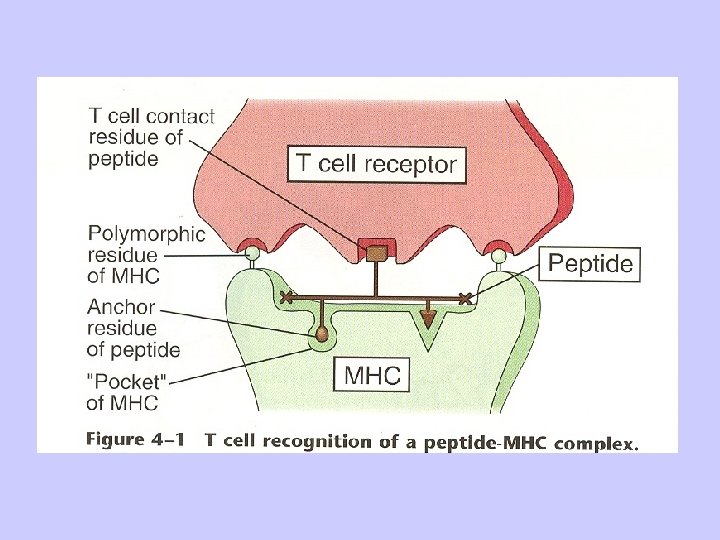
Peptide-binding region is a groove formed from the")
b. There are four separate regions: (1)Peptide-binding region is a groove formed from the alpha 1 and alpha 2 regions which interact to form a "floor" of an 8 -stranded, beta-pleated sheet with two opposite "walls" consisting of parallel strands of an a-helix. (alpha 1 and alpha 2 each contribute 4 strands of beta-pleated sheet and one alpha-helix). A peptide 8 -10 amino acids long sits in the groove. .
immunoglobulin-like region is composed of an alpha 3 segment that is highly conserved and")
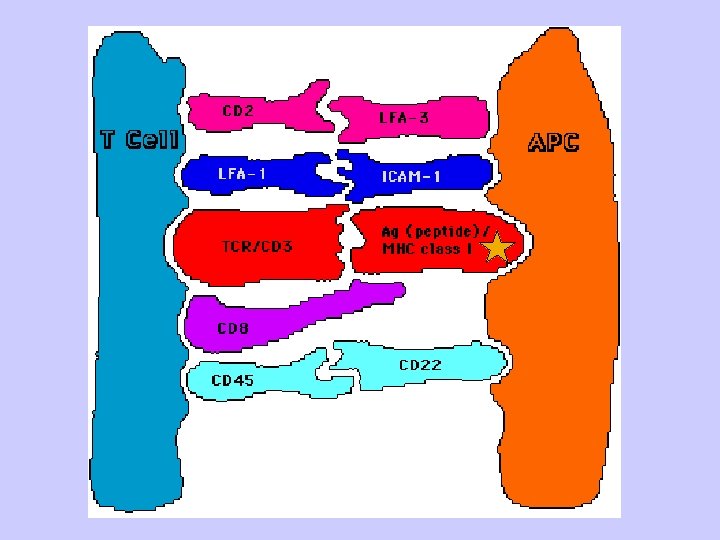
(2)immunoglobulin-like region is composed of an alpha 3 segment that is highly conserved and is homologous to Ig constant domains and non-covalently bound beta 2 microglobulin, an invariant molecule, also homologous to Ig constant domains. These two interact with alpha 1 and alpha 2 to maintain their proper conformation. The importance of the highly conserved region of alpha 3 is that CD 8, a molecule
transmembrane region is a stretch of ~25 hydrophobic amino acids. (4)cytoplasmic region is the")
(3)transmembrane region is a stretch of ~25 hydrophobic amino acids. (4)cytoplasmic region is the carboxy terminal 30 amino acids. Contains phosphorylation sites and provides binding sites for cytoskeletal elements.

Class I MHC

2. Class II MHC a. contains two non-covalently associated polypeptide chains, both MHC-encoded and polymorphic (1) alpha chain; 34 k. Da (2) beta chain; 28 k. Da
 peptide-binding region is formed by interaction of the alpha")
b. four separate regions (1) peptide-binding region is formed by interaction of the alpha 1 and beta 1 segments. There is a groove having a "floor" of 8 beta-pleated strands and two "walls" with alpha-helices; alpha 1 and beta 1 make equal contributions to this structure. The greatest polymorphic variability in the amino acids is in those facing the groove( b 1 region).
immunoglobulin-like region formed by alpha 2 and beta 2 is folded into Iglike domains.")
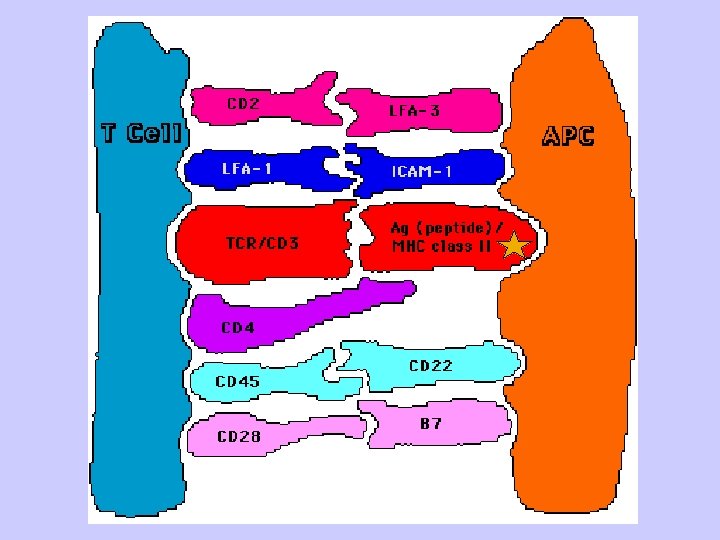
(2)immunoglobulin-like region formed by alpha 2 and beta 2 is folded into Iglike domains. These are largely nonpolymorphic. The correlation of CD 4 expression on helper T cells with a specific TCR for class II MHC molecules is due to binding of the CD 4 molecules to the Ig-like non-polymorphic beta 2 domain of the class II MHC molecules. (3, 4)transmembrane and cytoplasmic regions have proposed functions

Class II MHC

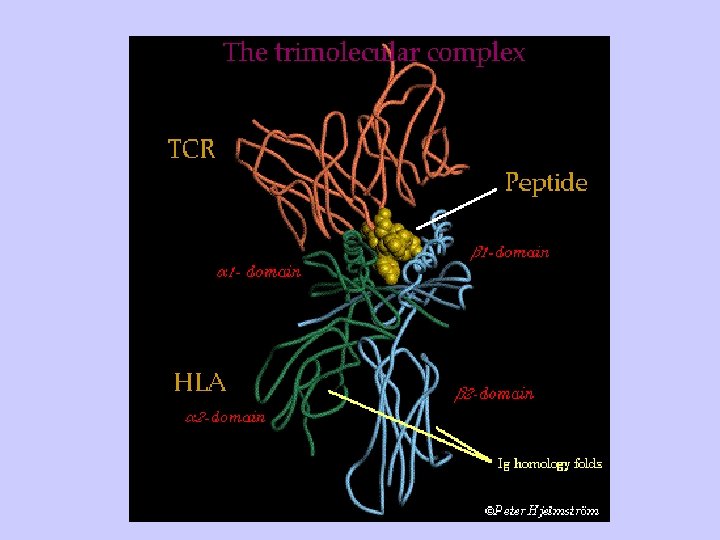
C. Similarities between class I and class II MHC molecules Despite differences in the two chain composition of class I and class II MHC molecules, they are quite similar structurally.

Expression of MHC molecules Cell Type MHC II T cells +++ + B cells Macrophages +++ +++ ++ Dendritic cells + + +X 10 Granulocytes Endothelium Hepatocytes Neurons ++ ++ + - (activated human T cells) - (all mouse T cells) + + +X 10 )- inducible( -

The polymorphic residues of Cl I &Cl II molecules: Cl I : located in the peptide-binding clefts and the a - helices around the clefts(a 1 & a 2). Cl II: show varying degrees of polymorphism in the a chain and usually much more in the b- chain (b 1).

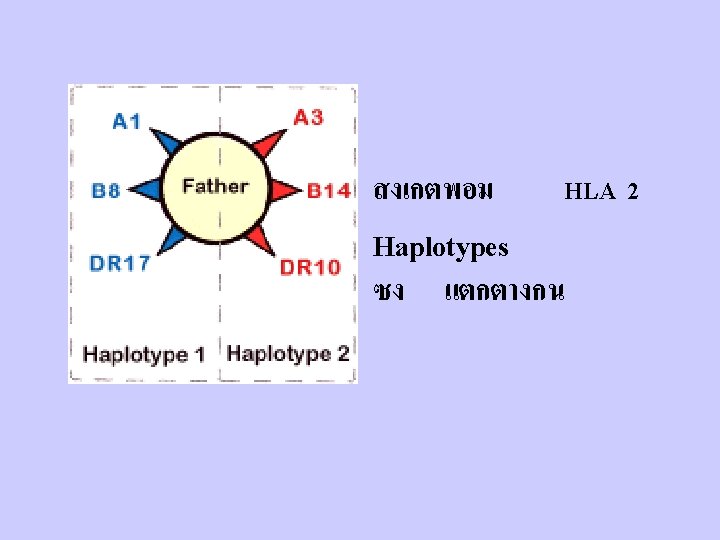
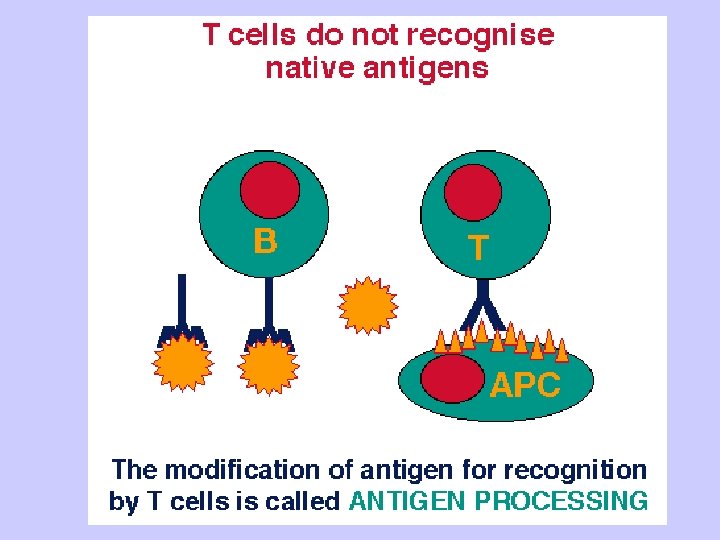
D. Important aspects of MHC 1. Because MHC molecules are membrane-associated and not soluble, T cells must make cell to cell contact with cells expressing MHC molecules. 2. Tc recognize class I MHC-peptide; Th recognize class II MHC-peptide. 3. There are many different MHC gene products for class I and class II in the human population (polymorphism)

4. Mature T cells respond to foreign antigens, but not self protein 5. Only a single binding site exists on a class I or class II MHC molecule; all peptides must bind to the same site. 6. The MHC polymorphism is determined only in the germline. There is no somatic DNA recombination that occurs for antibodies and for the TCR, so the MHC genes lack recombinational mechanisms for generating diversity. As a result, the affinity and selectivity of MHC molecules foreign proteins are considerably

7. Because each MHC molecule can bind many different peptides, the binding is said to be degenerate. Compare with the restricted binding of a hormone to a receptor, for example, where a difference in even one amino acid may impair binding. 8. Cytokines, especially interferon gamma (IFN-gamma), increase the level of expression of class I and class II MHC molecules 9. Alleles for MHC genes are co-


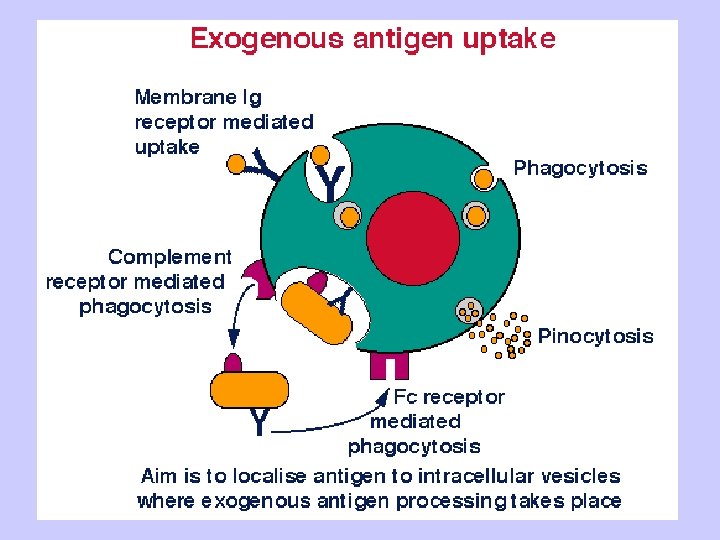
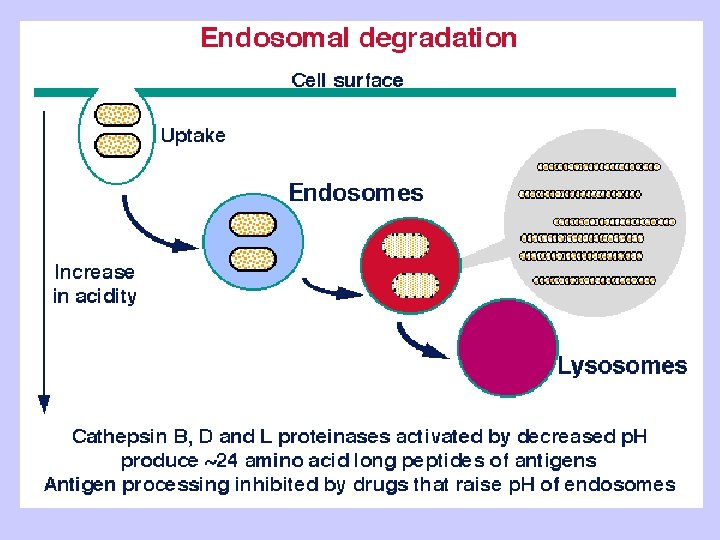
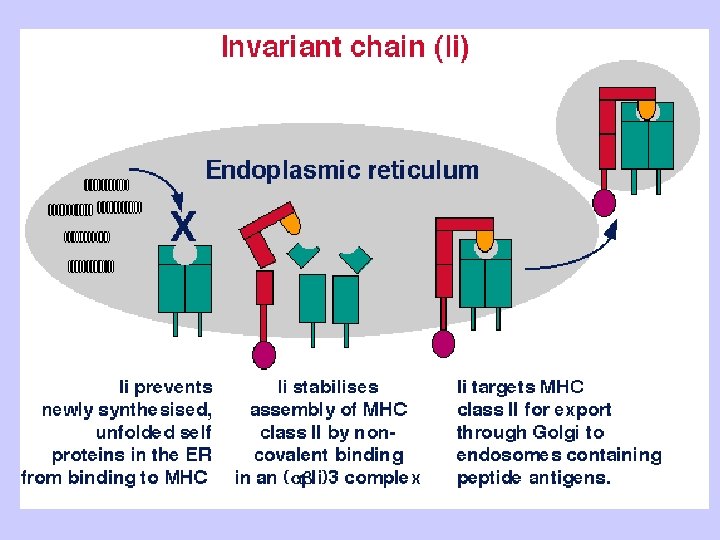
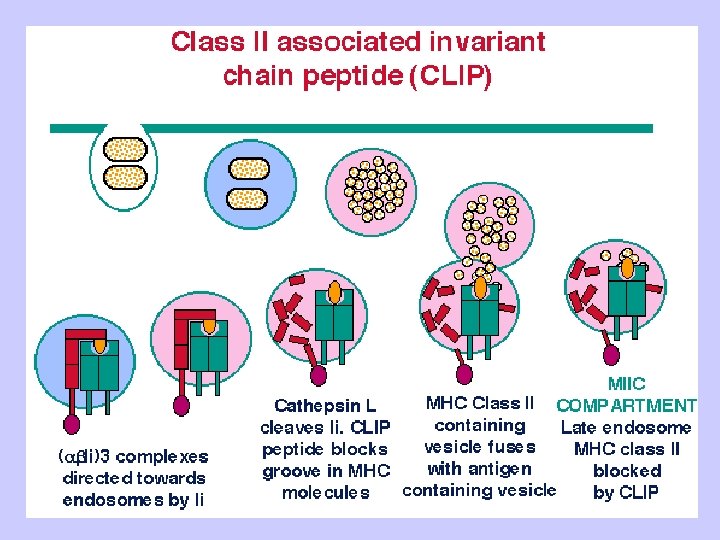
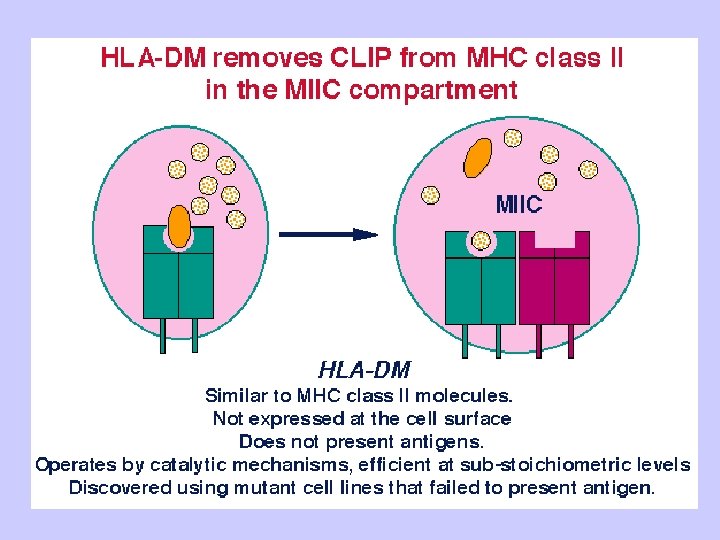
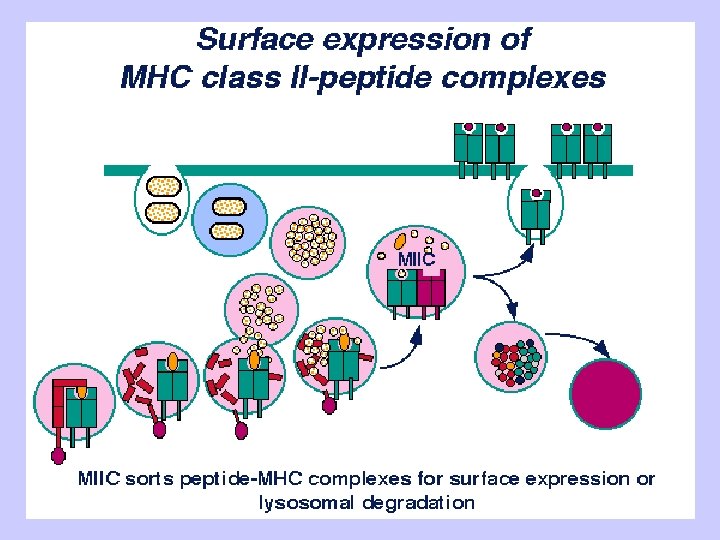
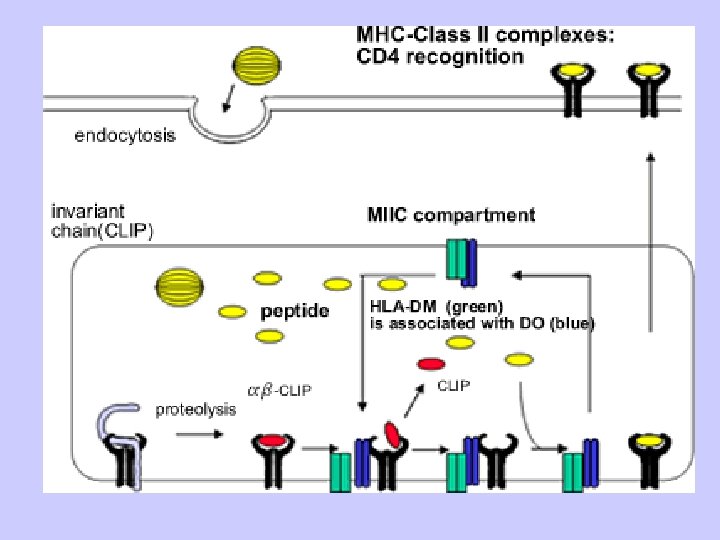
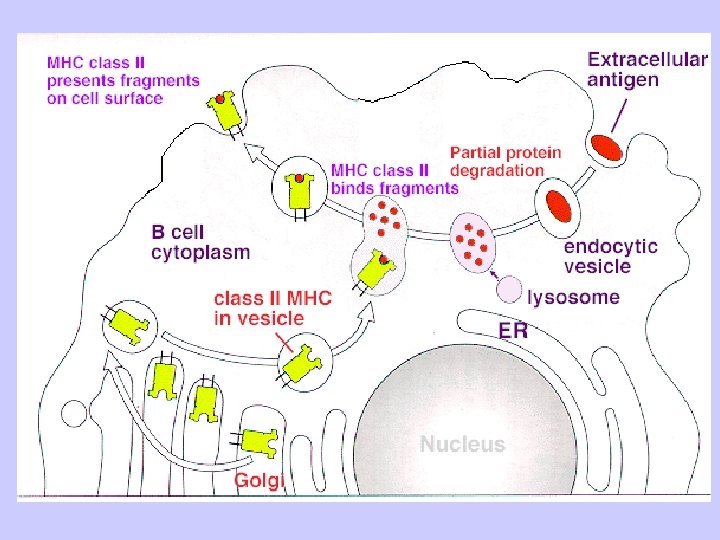
Exogenous antigen processing Uptake: Phagocytosis Pinocytosis Membrane Ig mediated uptake Degradation: Intracellular transport to vesicle system Proteolysis in acidified vesicles Complex formation: Invariant chain CLIP peptide HLA-DM Presentation: Transport of MHC-peptide complexes to cell surface Presenttation to CD 4+ T cells

8

PROTEOSOME
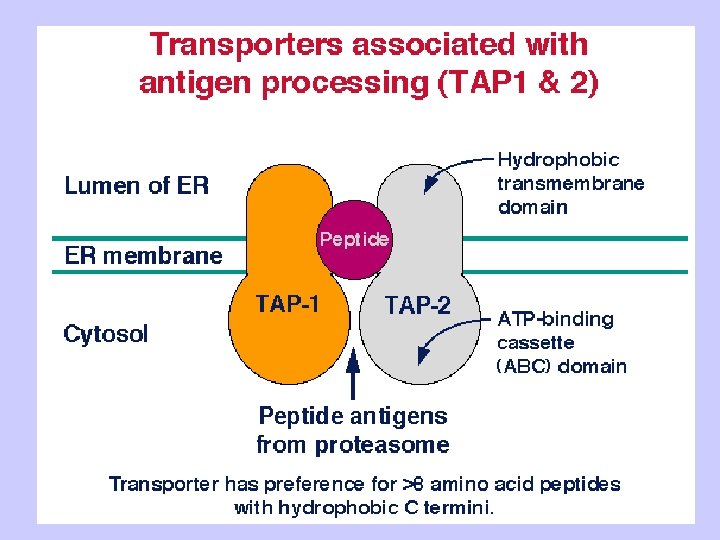

TAP: Transporter Associated with Antigen Processing
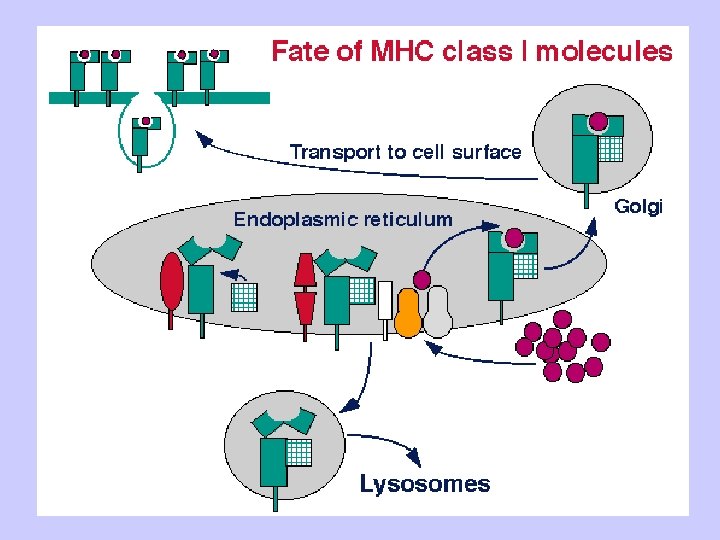
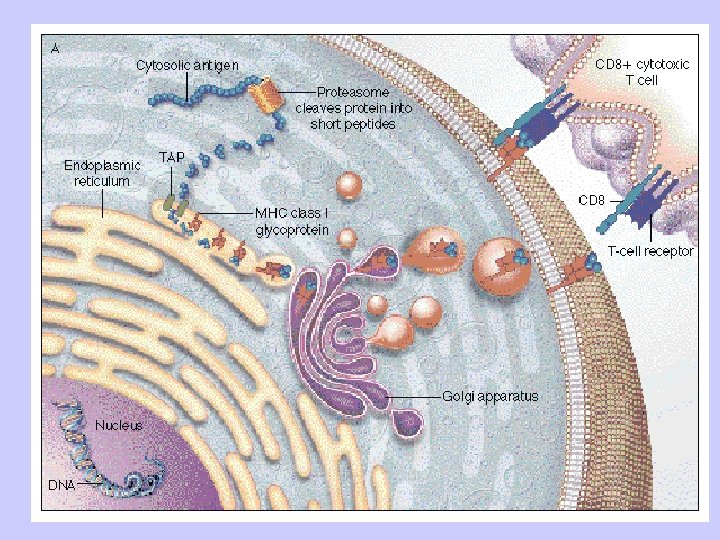
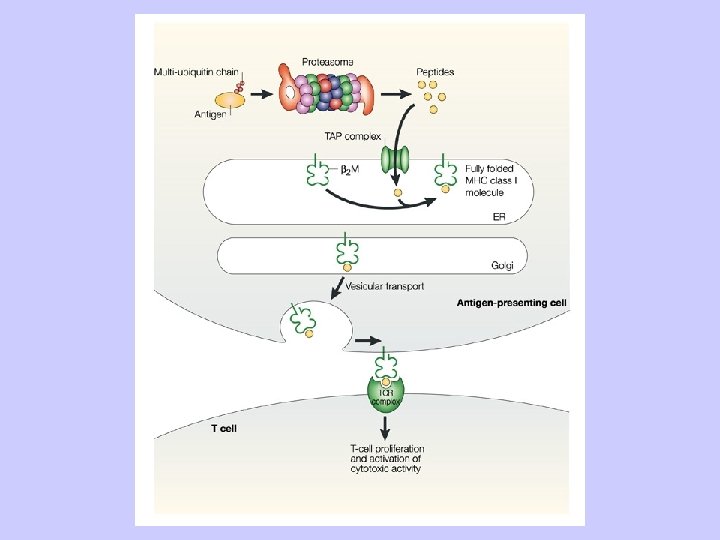

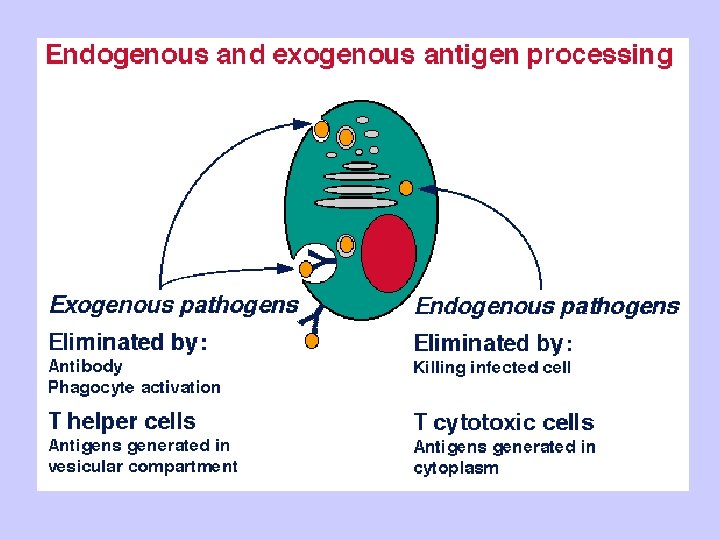
Antigen Processing and Presentation Class II MHC Endogenous pathway Exogenous pathway
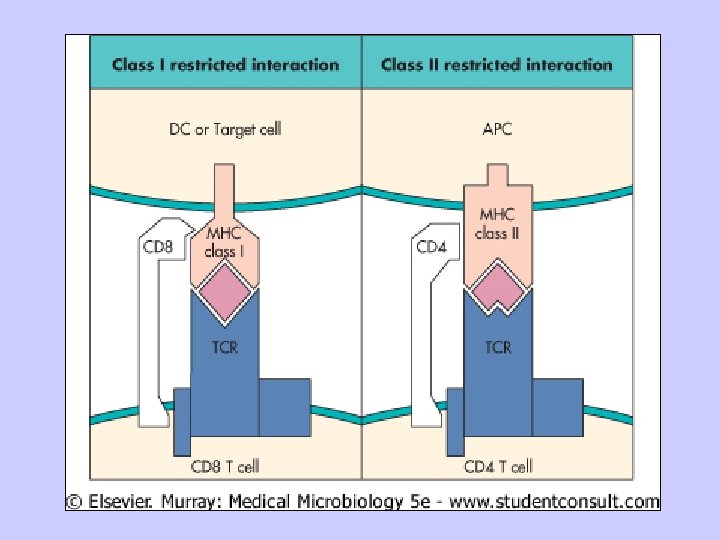

MHC restriction

Structure of the T cell Receptor • Heterodimer with one α and one β chain of roughly equal length • A short cytoplamic tail not capable of transducing an activation signal • A transmembrane region with hydrophobic amino acids 52

Structure of the T cell Receptor • Both α and β chains have a variable (V) and constant (C) region • V regions of the α and β chains contain hypervariable regions that determine the specificity for antigen 53

Structure of the T cell Receptor • Both α and β chains have a variable (V) and constant (C) region • V regions of the α and β chains contain hypervariable regions that determine the specificity for antigen • Each T cell bears TCRs of only one specificity (allelic exclusion) 54

Genetic Basis for Receptor Generation • Generation of a vast array of BCRs is accomplished by recombination of various V, D and J gene segments encoded in the germline • Generation of a vast array of TCRs is accomplished by similar mechanisms - TCR β chain genes have V, D and J gene segments - TCR α chain genes have V and J gene segments 55

Organization and Rearrangement of the T Cell Receptor Germline ß-Chain Gene L Vß 1 P L Vß 2 Dß 1 L Vßn P Jß 11 ----Jß 16 Cß 1 Dß 2 Jß 11 --------Jß 17 Cß 2 E P D-J rearrangement L P Vß 1 L Vß 2 L Vßn P Dß 1 Jß 15 Cß 1 Dß 2 Jß 11 --------Jß 17 Cß 2 P E V-D rearrangement L P LVß 2 Dß 1 Jß 15 Cß 1 Dß 2 Jß 11 --------Jß 17 Vß 1 P Cß 2 E Transcription Vß 2 Dß 1 Jß 15 Cß 1 RNA 56 DNA

γδ TCR • Small population of T cells express a TCR that contain γ and δ chains instead of α and β chains • The Gamma/Delta T cells predominate in the mucosal epithelia and have a repertoire biased toward certain bacterial and viral antigens • Genes for the δ chains have V, D and J gene segments; γ chains have V and J gene segments • Repertoire is limited • Gamma/Delta T cells can recognize antigen in an MHCindependent manner • Gamma/Delta T cells play a role in responses to certain viral and bacterial pathogens 57

Role of CD 3 Complex • CD 3 complex is necessary for cell surface expression of TCR during T cell development • CD 3 complex transduces signals to the interior of the cells following interaction of Ag with the TCR

Costimulation is Necessary for T Cell Activation • Engagement of TCR and Ag/MHC in the absence of costimulation can lead to anergy • Engagement of costimulatory molecules in the absenece of TCR engagement results in no response • Activation only occurs when both TCR and costimulatory molecules are engaged with their respective ligands • Downregulation occurs if CTLA-4 interacts with B 7 - CTLA-4 send inhibitory signal 59

Key Steps in T cell Activation • APC must process and present peptides to T cells • T cells must receive a costimulatory signal - Usually from CD 28/B 7 • Accessory adhesion molecules help to stabilize binding of T cell and APC - CD 4/MHC-class II or CD 8/MHC class I - LFA-1/ICAM-1 - CD 2/LFA-3 • Signal from cell surface is transmitted to nucleus - Second messengers • Cytokines produced to help drive cell division - IL-2 and others 60

- Slides: 63