Les cellules musculaires lisses Aptitude transformer lnergie chimique







")
 • Cellule fusiforme à un seul noyau")



 • Cytosquelette – filaments intermédiaires (desmine et")
 • Protéines contractiles – myosine II (SM-heavy")
 • Protéines associées aux filaments fins Tropomyosine")





 • Force")

 – Pas")













- Slides: 38
Les cellules musculaires lisses • Aptitude à transformer l’énergie chimique en énergie mécanique • Réalisation de contractions unidirectionnelles • Innervées par le système nerveux végétatif (pas de plaque motrice, pas de dégénérescence après dénervation)
Innervation des cellules musculaires lisses
Les cellules musculaires lisses • Transformation de l’énergie chimique en énergie mécanique • Réalisation de contractions unidirectionnelles • Innervées par le système nerveux végétatif (pas de plaque motrice, pas de dégénérescence après dénervation) • Dans certaines conditions, grande capacité à proliférer, migrer, synthétiser
Différents types de disposition des cellules musculaires lisses en couches isolées dans un organe regroupées en un muscle
Méthodes d’études du muscle lisse Organe isolé Culture de cellules
Culture de cellules musculaires lisses thymidine Étude de prolifération Étude de migration
Etude des canaux ioniques par patch-clamp
Mesure de la concentration de Ca intracellulaire par un indicateur fluorescent (fura-2)
Structure de la cellule musculaire lisse (1) • Cellule fusiforme à un seul noyau • Réticulum endoplasmique lisse et granulaire (synthèse d’élastine et de collagène de la matrice) • Membrane plasmique (sarcolemme) – caveolae – plaques d ’adhérence
La cellule musculaire lisse MEx 5500 MEx 32400 Le sarcolemme présente de nombreux caveolae et des plaques d’adhérence.
Rôle des caveolae dans le couplage excitation-contraction
Sites d’attachement des filaments contractiles Filaments contractiles d’actine et myosine Filaments intermédiaires Filament d’actine Corps denses Plaques d’adhérence intégrine vitronectine Plaque d’adhérence, Zones d’ancrage de filaments d’actine à la matrice extracellulaire par l’intermédiaire des intégrines
Structure de la cellule musculaire lisse (2) • Cytosquelette – filaments intermédiaires (desmine et vimentine) – filaments d’actine non musculaire et smoothelin – corps denses (actine non musculaire et aactinine)
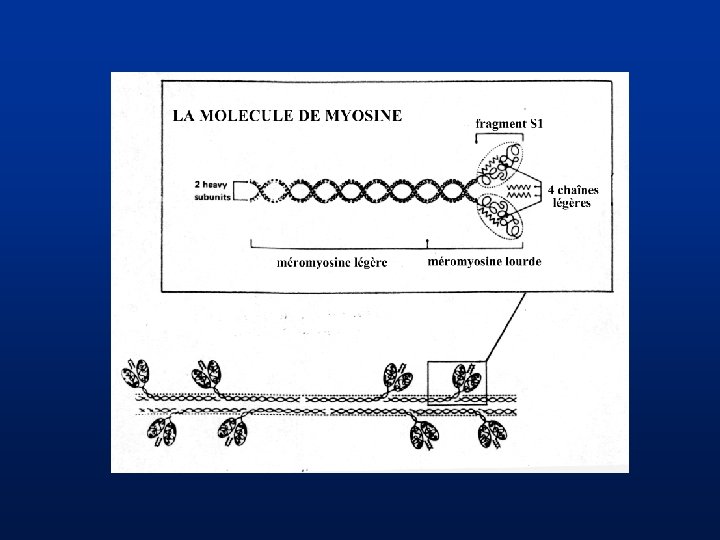
Structure de la cellule musculaire lisse (3) • Protéines contractiles – myosine II (SM-heavy chain 1 et 2 codées par le même gène, chaînes légères régulatrices et essentielles) – SM actine (2 isoformes, a et g)
Structure de la cellule musculaire lisse (4) • Protéines associées aux filaments fins Tropomyosine (plusieurs gènes et plusieurs isoformes par gène) caldesmone ( 2 isoformes codées par un même gène, inhibition de l’interaction actine-myosine levée par sa phosphorylation) calponine (3 isoformes produites par des gènes différents dont 1 spec du muscle lisse, inhibition de l’actine myosine ATPase) SM 22
Formation cyclique des ponts actine myosine
Activation des protéines contractiles Ca 2+ + Ca. M Ca 2+ 4. Ca. M + MLCK (inactive) Ca 2+ 4. Ca. M + MLCK (active) ATP ADP MP L’intensité de la force proportionnelle à la concentration de Ca 2+ et à la phosphorylation de la chaîne légère régulatrice A AMP Contraction phasique
La déphosphorylation de la chaîne légère est catalysée par une phosphatase dont dépend la vitesse de relaxation Ca 2+ + Ca. M Ca 2+ 4. Ca. M + MLCK (inactive) Ca 2+ 4. Ca. M + MLCK (active) ATP A ADP MP M Pi Pi AM A phosphatase AMP Contraction phasique
Changement d’affinité des protéines contractiles pour le Ca 2+ Tout facteur capable de modifier l’activité de la MLCK ou de la phosphatase peut provoquer une variation de la contraction indépendamment de la concentration de Ca 2+
Activation des protéines contractiles Ca 2+ + Ca. M Ca 2+ 4. Ca. M + MLCK (inactive) Ca 2+ 4. Ca. M + MLCK (active) ATP ADP MP Pi Pi AM Contraction verrouillée A phosphatase AMP Contraction phasique
Energétique de la contraction du muscle lisse (comparaison avec le muscle strié) • Force développée (nombre de ponts) aussi grande • Raccourcissement peut être plus grand • Vitesse de raccourcissement (ATPase de la myosine) plus lente et consommation d’énergie plus faible (selon isoformes rapides ou lentes, muscles phasiques et toniques) • Contraction maintenue pendant des heures au prix d’une faible dépense d’énergie (contraction verrouillée ou « latch contraction »
Une particularité du muscle lisse Capacité à réorganiser ses filaments contractiles en réponse à un changement de longueur ou de tension ØForce maximale générée relativement indépendante de la longueur de repos
Deux types de muscle lisse • Muscle lisse multi-unitaire (muscle pilo-moteur, ciliaire) – Pas de potentiels d’action spontanés – Pas de propagation de potentiels d’action de cellules en cellules • Muscle lisse unitaire (muscle lisse vasculaire, du tube digestif) – Potentiels d’action spontanés qui se propagent grâce aux jonctions communicantes
Dépolarisations spontanées de cellules musculaires lisses 2 sec
Le muscle lisse des voies aériennes Électriquement quiescent mais il existe des jonctions communicantes entre cellules
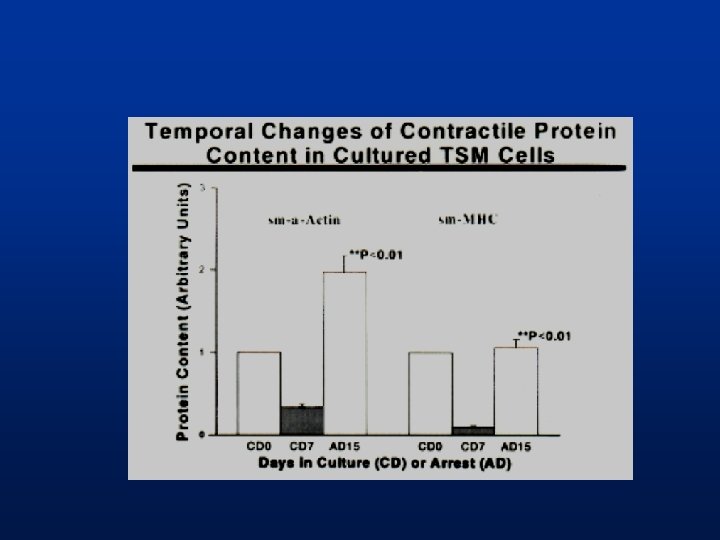
Phénotype des cellules musculaires lisses en culture
Différents phénotypes de cellules musculaires lisses dans la paroi de l’artère carotide de chien
Contrôle du muscle lisse par les messagers extracellulaires • Des messagers extracellulaires contrôlent la contraction du muscle lisse grâce à une interaction avec des récepteurs spécifiques présents sur la membrane cellulaire • La transduction du message recouvre l’ensemble des phénomènes qui traduisent l’information extracellulaire en variation de la concentration de Ca 2+ cytosolique
Contrôle du muscle lisse par les substances libérées dans le milieu extracellulaire (hormones, autacoïdes, neuromédiateurs) • Couplage électromécanique : variation de potentiel localisée avec ou sans potentiel d’action • Couplage pharmacomécanique : absence de variation de potentiel
Couplage électromécanique Couplage pharmacomécanique Ca 2+ R b bg GTP Activation d’un canal ionique Couplage électromécanique E a Activation d’enzyme Couplage pharmacomécanique
Caractéristiques du potentiel d’action du muscle lisse • Potentiel de repos (- 50 à - 60 mv) • Durée du potentiel d’action (10 à 50 ms voire 300 ms) • Potentiel d’action lié à l’entrée d’ions Ca 2+ essentiellement
Voltage dependent 2+ Ca channels Ca 2+ 2 K+ 3 Na+ Ca 2+ 2 H+ Ca 2+ ATP dépolarisation Ca 2+ K+ Receptor operated channels
Protéine Rho et sensibilité au Ca 2+ des protéines contractiles agoniste G 12/13 Gq DAG PLC IP 3 Rho-GTP Ca 2+ Rho-kinase CPI-17 M 20 Ca. M MLCK CPI-17 -P MLC P MBS Cat Ca 2+ M 20 MBS MLC-P Cat Phosphatase
Cl- Ca 2+ K+ Ca 2+ R 1 ROC SOC PI G DAG IP 3 rya Ca 2+ Déplétion en Ca 2+ ATPase PKG Myosine MLCK Myosine P phosphatase DAG PKC Rho-K AMPc PKA GMPc contraction G G R 2 R 3
La contraction est inhibée • En cas de couplage électromécanique par l’absence de Ca 2+ extracellulaire ou la DHP, inhibiteur des canaux calciques dépendant du voltage • En cas de couplage pharmacomécanique, par la thapsigargine qui déplète le réticulum en Ca 2+
Rôle du Ca 2+ dans la prolifératon des CML Ca 2+noyau Ca 2+cytosol Ca 2+ - calmodulinedependent kinases prolifération Ca 2+RS