LECTURE PRESENTATIONS For CAMPBELL BIOLOGY NINTH EDITION Jane

LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 12 & 18 Control of DNA Lectures by Erin Barley Kathleen Fitzpatrick © 2011 Pearson Education, Inc.

Eukaryotic Cell Cycle Control • the frequency of cell division varies with the type of cell – differences result from regulation at the molecular level – cancer cells manage to escape the usual controls on the cell cycle • cell cycle is driven by specific chemical signals present in the cytoplasm EXPERIMENT Experiment 1 S G 1 S S Experiment 2 M G 1 RESULTS When a cell in the S phase was fused with a cell in G 1, the G 1 nucleus immediately entered the S phase—DNA was synthesized. M M When a cell in the M phase was fused with a cell in G 1, the G 1 nucleus immediately began mitosis—a spindle formed and chromatin condensed, even though the chromosome had not been duplicated.
 phase")
Phases of the Cell Cycle • consists of two phases – Mitotic (M) phase = mitosis and cytokinesis) – Interphase = cell growth and copying of chromosomes in preparation for cell division • Interphase - about 90% of the cell cycle – – can be divided into sub-phases G 1 phase -“first gap” S phase “synthesis” G 2 phase - “second gap”

Some terms to know -parent cell = cell about to undergo division -daughter cell = cell that results from either mitosis or meiosis -somatic cell = any cell within the body other than an egg or sperm -somatic cell has two complete sets of chromosomes -one set is called the haploid number of chromosomes (n) -therefore the cell is said to be diploid (2 n) e. g. humans n = 23 (2 n = 46) -germ cell or gamete = sex cell -gamete has only one set of chromosomes and is haploid

Cellular Organization of the Genetic Material • all the DNA in a cell constitutes the cell’s genome • REMINDER: eukaryotic chromosomes consist of chromatin, a complex of DNA and protein that condenses during cell division • in most cells - DNA molecules in a cell are condensed and packaged into chromosomes • prokaryotics have a single chromosome called a genophore • eukaryotic cells posses number of chromosomes 20 m every eukaryotic species has a characteristic number of chromosomes in each cell nucleus e. g. humans – n=23 e. g. drosophila – n=2 e. g. dog – n=39

• when not dividing – much of the eukaryotic DNA is in its loosest formation = chromatin – allows access to the machinery for DNA replication and transcription • in preparation for cell division - DNA is replicated and condenses into chromosomes • chromosome = organized structure of DNA and protein – chroma = color – soma = body • the building material of a chromosome is chromatin • each duplicated chromosome is made of two sister chromatids = joined copies of the original chromosome – these chromatids will separate during cell division and be partitioned into each daughter cell • chromatids are joined by a structure called a centromere Sister chromatids Centromere 0. 5 m

Centromere • condensed region within the chromosome • responsible for the accurate segregation of sister chromatids during mitosis & meiosis • shared by sister chromatids during mitosis • site of the centromere where spindle microtubules attach – area of DNA and protein = kinetochore Sister chromatids Centromere 0. 5 m

Kinetochore a structure of DNA and proteins located in the centromere for the attachment of the chromosome to the spindle during mitosis and meiosis one MT attaches to one kinetochore on one chromatid a 2 nd MT attaches to the kinetochore on the other chromatid attachment of MTs to the plates of the kinetochore results in movement toward the poles • a “tug of war” results – chromosomes move back and forth and eventually settle in the metaphase plate • • • Chromatid Outer Plate Microtubules Kinetochore Microtubules Inner Plate

Control of DNA Structure: Condensation of Chromatin • nucleosome = DNA helix wrapped around a histone protein core – responsible for organizing the DNA as chromatin • • other proteins are involved organizing chromatin as chromosomes non-histone proteins in the nucleus form a chromosome scaffold 300 nm fiber specific sequences in the chromosome called scaffold-associated regions or SARs interact with the scaffold to create the 300 nm fiber next level of “packing” is a 700 nm fiber – not much known about this http: //www. ndsu. edu/pubweb/~mcclean/plsc 431/eukarychrom/eukaryo 3. htm chromosome scaffold

Chromosome and Chromosomes: Confusion!!! • prior to cell division – the duplicated chromatin condenses into its most dense form = chromosome – two sister chromatids joined by a centromere – typically called a duplicated chromosome • during cell division - the two sister chromatids separate • once separated - the chromatids are still called chromosomes DNA condensation animation http: //www. biostudio. com/demo_fr eeman_dna_coiling. htm

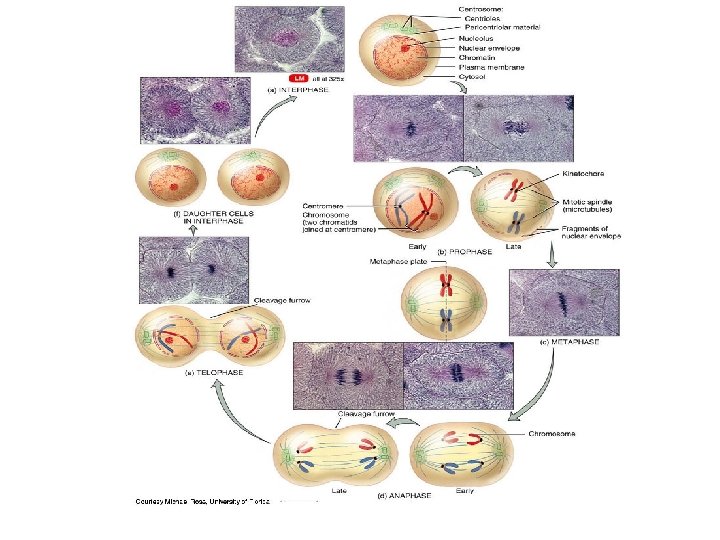
• Mitosis is conventionally divided into five phases – – – Prophase Prometaphase Metaphase Anaphase Telophase • Cytokinesis overlaps the latter stages of mitosis
 Nucleolus Chromatin (duplicated) Nuclear")
10 m G 2 of Interphase Centrosomes (with centriole pairs) Nucleolus Chromatin (duplicated) Nuclear envelope Plasma membrane Prophase Early mitotic spindle Aster Centromere Chromosome, consisting of two sister chromatids Prometaphase Fragments of nuclear envelope Kinetochore Metaphase Nonkinetochore microtubules Kinetochore microtubule Anaphase Metaphase plate Spindle Centrosome at one spindle pole Telophase and Cytokinesis Cleavage furrow Daughter chromosomes Nuclear envelope forming Nucleolus forming

Mitosis 1. http: //www. loci. wisc. edu/outreach/bioclips/CDBio. html Prophase: prior to prophase, the replicated DNA is beginning to condense into sister chromatids joined at the centromere (duplicated) chromosome 1. the centrioles (replicated at G 2) move apart from each other 2. the spindle forms between the centrioles (made of microtubules) 3. the kinetochore forms and the condensing duplicated chromosomes attach to the spindle 4. the nucleoli disappear Spindle – structure that includes the two centrioles, two asters and the spindle microtubules than span the cell Aster – a radial array of short MTs extending from the centrioles

2. Prometaphase: used to be known as “late prophase” 1. the nuclear envelope fragments – allows growth of spindle into region where chromosomes are located 2. the DNA of the duplicated chromosomes becomes even more condensed 3. some chromosomes attach to spindle via kinetochore = kinetochore microtubules 4. non-kinetochore microtubules begin to form and grow towards opposite pole Prometaphase Fragments of nuclear envelope Kinetochore Nonkinetochore microtubules Kinetochore microtubule

3. Metaphase: centrioles are at opposite ends of the cell and the spindle is complete 1. the chromosomes move and line up along a central zone= metaphase plate -the tug of war at pro-metaphase eventually positions the chromosomes midway alone the length of the cell 2. non-kinetochore MTs interact with the opposite pole & the aster MTs make contact with the plasma membrane – the spindle is now complete 10 m 3 Metaphase

4. Anaphase: shortest of the mitotic phases 1. the chromatid pairs separate into daughter chromosomes 2. one chromatid/chromosome moves toward one centriole of the cell, the other the opposite -pulled apart by the action of the spindle – the kinetochore MTs begin to shorten -PLUS non-kinetochore MTs grow – this elongates the cell ** At the end of this phase – each end of the cell has equivalent numbers of chromosomes – same number as the parent cell **the sister chromatids separate because of enzymatic activity -an enzyme called separase cleaves a protein known as cohesin (protein in the centromere that holds the sister chromatids together) -separates the sister chromatids

4. Telophase: reverse of Prophase 1. nuclear envelope reforms – two daughter nuclei result -part of the new nuclear membrane is recycled from the old fragments, other parts are made new by the cell 2. the nucleoli reappear 3. the spindle disappears as the MTs depolymerize 4. daughter chromosomes uncoil ** Cytokinesis starts during late anaphase and is well underway during telophase
 Cleavage of an animal cell (SEM) Cytokinesis: division of cytoplasm -separates the parent")
(a) Cleavage of an animal cell (SEM) Cytokinesis: division of cytoplasm -separates the parent into two daughter cells -differs in animal cells and plant cells Animal cell Cytokinesis: results from cleavage -pinches into two daughters -actin filaments assemble to form a contractile ring along the equator of the cell -actin interacts with myosin proteins – causes the ring to contract -actin-myosin interaction first forms a “cleavage furrow” - slight indentation around the circumference of the cell -continued interaction divides the cell by a “purse string” mechanism Cleavage furrow Contractile ring of microfilaments 100 m Daughter cells

Plant cell Cytokinesis: No cleavage furrow possible -vesicles bud from the Golgi apparatus and migrate to the middle of the cell -vesicles coalesce to produce a cell plate -other vesicles fuse to the plate bringing in new building materials -cell plate grows and eventually splits the cell into two daughter cells Cell plate 10 m (b) Cell plate formation in a plant cell (TEM) Vesicles forming cell plate 5 Telophase Wall of parent cell Cell plate 1 m New cell wall Daughter cells

Binary Fission in Bacteria • bacteria and archaea reproduce by binary fission – the chromosome replicates and the two daughter chromosomes actively move apart – the plasma membrane pinches inward, dividing the cell into two – BUT there is no ordered segregation of the duplicated chromosome – duplicate it and divide it into two cells Origin of replication E. coli cell 1 Chromosome replication begins. 2 Replication continues. 3 Replication finishes. 4 Two daughter cells result. Cell wall Plasma membrane Bacterial chromosome Two copies of origin Origin

The Evolution of Mitosis • mitosis probably evolved from binary fission • certain protists exhibit types of cell division that seem intermediate between binary fission and mitosis (a) Bacterial chromosome Chromosomes (b) Dinoflagellates Microtubules Intact nuclear envelope Kinetochore microtubule (c)Diatoms and some yeasts Intact nuclear envelope Kinetochore microtubule (d) Most eukaryotes Fragments of nuclear envelope

Phases of the Cell Cycle – G 1 phase - time in phase depends on species • • normal cell functions growth in size m. RNA and protein synthesis in preparation for S phase critical phase in which cell commits to division or leaves the cell cycle to enter into a dormancy phase (G 0) – S phase - 6 to 8 hours • synthesis of histone proteins & DNA replication – G 2 phase – 2 to 5 hours • rapid cell growth – may function to simply control cell size • protein synthesis in preparation for M phase • duplication of the centrioles/centrosomes http: //www. wisconline. com/objects/in dex. asp? obj. ID=AP 136 04

Cell Cycle Checkpoints • interphase not only allows the cell to perform its normal functions but also allows the cell to check whether it is ready to enter mitosis • cell cycle is controlled by a control system that coordinates and triggers key events in the cell cycle • progression through the cell cycle requires a combination of internal and external signals – these signals control whether the cell is ready to continue on into the S and M phases • so the control system of the cell cycle monitors these signals and determines whether to proceed through the cell cycle • there are specific points along the cell cycle where “decisions” are made by this control system = CHECKPOINTS G checkpoint • Checkpoint = control point where “stop” and “go-ahead” signals regulate the cell cycle 1 Control system S G 1 M G 2 M checkpoint G 2 checkpoint

Cell Cycle Checkpoints • major checkpoints – G 1, G 2 and M – G 1 checkpoint – G 1/S progression through a point called the restriction point or START point of the cell cycle – G 2 checkpoint – G 2/M progression which will lead to the start of mitosis and chromosome alignment – M checkpoint – Metaphase to Anaphase transition where chromatid separation occurs G 1 checkpoint Control system G 1 M G 2 M checkpoint G 2 checkpoint S

Cell Cycle Checkpoints • for many cells, the G 1/S checkpoint seems to be the most important – if a cell receives a go-ahead signal at this G 1/S checkpoint will usually complete the S, G 2, and M phases and divide – many texts call this checkpoint the Start (yeasts) or Restriction point (mammalian cells) • if the cell does not receive the go-ahead signal - will exit the cycle, switching into a non-dividing state called the G 0 phase • most cells in the body are in the G 0 phase and remain there • some cells have the ability to leave G 0 and re-enter the cell cycle G 0 G 1 checkpoint G 1 (a) Cell receives a go-ahead signal. G 1 (b) Cell does not receive a go-ahead signal.

IN ANIMAL CELLS A “GO-AHEAD” SIGNAL MUST BE PRODUCED TO OVERRIDE A BUILT IN “STOP” SIGNAL

Cell Cycle Control System • the proteins of this system evolved over a billion years ago • so well conserved in eukaryotes – take from human control cells and put into yeast cells – they work!! • much of the early research done – been done in yeast – search for mutations in genes that encode critical parts of the cell cycle control system = cell-division-cycle genes or cdc genes – many mutations cause the cell cycle to arrest a specific points – such as checkpoints • additional work done in frog eggs and in mammalian cell cultures – e. g. immortalized mammalian cell lines

The Cell Cycle Control System: Cyclins and Cyclin. Dependent Kinases • master controllers of the cell cycle are heterodimeric protein kinases: cyclins and cyclin-dependent kinases (CDKs) – cyclins named for the cyclical changes in their concentration through the cell cycle • only made during the stage of the cell cycle they control – CDKs named because their phosphorylation activities requiring their binding to their “partner” cyclin • CDK levels remain constant throughout the cell cycle • it is the cyclins that cycle up and down – CDKs cannot work as kinases unless they are bound to their partner cyclin

The Cell Cycle Control System: Cyclins and Cyclin. Dependent Kinases – cyclin/CDKs regulate the activities of multiple proteins by the cdk phosphorylating them – Cyclin/CDK partners are involved in: • 1. entry to the cell cycle (i. e. G 1 phase) • 2. DNA replication (i. e. S phase) • 3. segregation of chromosomes during mitosis (i. e. M phase) – mistakes only happen once every 104 105 divisions phosphorylation can result in activation of the target protein or inhibition of the target protein (all depends on the target protein)

The Cell Cycle Control System: Cyclins and Cyclin. Dependent Kinases • CDK activity fluctuates during the cell cycle – CDK proteins are expressed first – CDK activity is controlled by an array of proteins - including the cyclin – cyclical changes in CDK activities leads to cyclical changes in the phosphorylation of their target proteins – CDKs phosphorylate key proteins responsible for passing through each checkpoint • e. g. phosphorylation of proteins that make up a pre-initiation complex - this results in the initiation of DNA replication

The Cell Cycle Control System: Cyclins and Cyclin. Dependent Kinases • eukaryotic cells have four classes of cyclins – each act at a specific stage of the cell cycle – eukaryotic cells require three of these classes for their cell cycle • 1. G 1/S cyclins • 2. S-cyclins • 3. M-cyclins

The Cell Cycle Control System: Cyclins and Cyclin. Dependent Kinases • 1. G 1/S cyclins – active in late G 1 – trigger the progression through the G 1 restriction point – cyclin E; cdk 2 • 2. S-cyclins – activate cdks that help stimulation chromosome duplication – levels stay high until M phase - since they help initiate mitosis – cyclin A; cdk 2 – E. g. cdk 2 activates the helicase • 3. M-cyclins – activate the cdks that stimulate progression through the G 2 checkpoint and into Mitosis – cyclin B; cdk 1

Cyclins and Cyclin-Dependent Kinases – no you don’t have to know these for your exam!!!

• • • work in frog eggs identified a cdk-cyclin complex that triggered the cell’s passage past the G 2 checkpoint into the M phase called MPF (maturation-promoting factor) • also called Mitosis-promoting factor – actually - cyclin B and cdk 1 cyclin B combines with cdk 1 in G 2 to produce MPF – cyclin B levels peak at M phase – MPF activity peaks at M phase when enough MPF is made – the cell passes the G 2 checkpoint and enters Mitosis during anaphase cyclin B is degraded and cdk 1 activity starts to fall M phase stops and G 1 begins – new cell cycle M G 1 S G 2 MPF activity Cyclin concentration Time (a) Fluctuation of MPF activity and cyclin concentration during the cell cycle M G 1

Cyclins and CDKs in action • work in frog eggs identified a cdk-cyclin complex that triggered the cell’s passage past the G 2 checkpoint into the M phase • called MPF (maturation-promoting factor) • also called Mitosis-promoting factor – cyclin B and CDK 1 • now known to act in all eukaryotic mitotic cells M G 1 S G 2 M G 1 S MPF activity Cyclin concentration Time (a) Fluctuation of MPF activity and cyclin concentration during the cell cycle G 2 M G 1

Cyclins and CDKs in action • cyclin B synthesis begins in late S phase and increases through G 2 and into M • cyclin B combines with CDK 1 in G 2 to produce MPF • when enough MPF is made – the cell passes the G 2 checkpoint and enters Mitosis • MPF promotes Mitosis via CDK 1 phosphorylating its target proteins – e. g. nuclear lamina proteins nuclear envelope fragmentation – e. g. chromosome condensing

Regulation of CDK activity M • M phase stops and G 1 begins G 1 S G 2 M G 1 MPF activity Cyclin concentration 1 Time (a) Fluctuation of MPF activity and cyclin concentration during the cell cycle S Cdk Cyclin is degraded M 2 Degraded cyclin G – the cdk 1 part of the MPF actually degrades its partner cyclin – the cdk 1 is “recycled” for future cycles M G • want to control the CDK? control the expression of the cyclin!!! • the most important regulatory control of a cyclin is its degradation • e. g. during anaphase cyclin B becomes degraded and CDK 1 activity starts to fall G 1 S G 2 Cdk checkpoint MPF Cyclin (b) Molecular mechanisms that help regulate the cell cycle

Degradation of cyclins and cell cycle control • activation of specific cyclin-cdk complexes also drive progression through the G 1 and G 2 checkpoints • progression through the M checkpoint requires the degradation of proteins • e. g. degradation of cyclin B • one key regulator of cyclin degradation is the anaphasepromoting complex or APC • mitotic cdk/cyclin complexes induce chromosome condensing, breakdown of the nuclear envelope, assembly of the spindle and the alignment of chromosomes – this gets you up to Anaphase • once this happens – the sister chromatids must separate at Anaphase • the APC controls this

Degradation of cyclins and cell cycle control cessation of mitosis • mitotic cdk/cyclins activate the APC (via phosphorylation) • the APC directs the degradation of proteins that act as anaphase inhibitors in the cell • allows the onset of anaphase cyclin/cdk activates APC degradation of inhibitors anaphase

Degradation of cyclins and cell cycle control cessation of mitosis • BUT as mitosis proceeds – the APC also directs the degradation of the cdk/cyclin complex • increasing APC levels act through negative feedback • the drop in mitotic cdk/cyclin complexes now allows for the condensing of chromosomes etc…. . and the cessation of mitosis cyclin/cdk promotes degradation activates APC degradation of inhibitors anaphase

Stop and Go Signs: Internal and External Signals at the Checkpoints • there is a link between what is happening inside and outside the cell with the activity of cdk/cyclins • in other words – internal and external signals exert control over cdk/cyclins and the cell cycle • internal signal – e. g. kinetochores not attached to spindle microtubules send a molecular signal that delays anaphase – all chromosomes must be attached to the spindle in order to eventually activate and enzyme called separase – separase promotes chromatid separation during anaphase

Stop and Go Signs: Internal and External Signals at the Checkpoints • some external signals are growth factors – proteins released by certain cells that stimulate other cells to divide = mitogen – more than 50 growth factors identified in eukaryotic cells – some GFs are made by several types of cells – others are quite cell-specific • another external signal - density-dependent inhibition – crowded cells stop dividing • another signal - anchorage dependence – cells must be attached to a substratum in order to divide

Scalpels 1 A sample of human connective tissue is cut up into small pieces. 2 Enzymes digest the extracellular matrix, resulting in a suspension of free fibroblasts. External Cell-Cycle Signals Petri dish 3 Cells are transferred to culture vessels. Without PDGF 4 PDGF is added to half the vessels. 10 m With PDGF platelet-derived growth factor (PDGF) stimulates the division of human fibroblast cells in culture – bind to PDGF receptors on target cells – initiate the progression through the G 1 phase into M • • GFs like PDGF often initiate a series of internal signaling events that activate a class of kinases called G 1 kinases -G 1 kinases then exert control over the G 1/S cyclins that initiate entry into the cell cycle

Loss of Cell Cycle Controls in Cancer Cells • cancer cells exhibit neither density-dependent inhibition nor anchorage dependence • Cancer = known medically as a malignant neoplasm • cells with upregulated cell growth because they do not have a normally controlled cell cycle • uncontrolled division gives rise to cells that can invade surrounding tissues and travel through the lymphatic and circulatory systems to invade other tissues at a distance = malignant cancer • if abnormal cells remain only at the original site - the lump is called a benign tumor Lymph vessel Blood vessel Tumor Cancer cell Glandular tissue 1 A tumor grows from a single cancer cell. Metastatic tumor 2 Cancer cells invade neighboring tissue. 3 Cancer cells spread through lymph and blood vessels to other parts of the body. 4 Cancer cells may survive and establish a new tumor in another part of the body.

Causes of Cancer • over 200 cancers that affect humans – Thyroid cancer – 94% five year survival rate – Skin cancer – 81% Mutagen – agent that causes a – Prostate cancer – 74% mutation in the DNA – Breast cancer – 76% Carcinogen – mutagen that causes – Stomach cancer – 17% cancer – Pancreatic – 1% • numerous and diverse causes – 90 -95% cases are attributed to environmental causes • Environmental causes = not due to genetic, inherited factors – Tobacco – 25 -30% of all cancers – Diet & obesity – 30 -35% • negative effects on the immune system and endocrine system • e. g. high salt diet – tied to increased gastric cancer – Infective agents (virus, bacteria) – 15 -20% – Radiation – up to 10%

Mutagens • mutagen = agent that causes a mutation in the DNA • mutations can arise in a number of ways – 1. errors in replication – 2. errors in genetic recombination (during meiosis) • mutagens can be both physical and chemical agents – physical – X-rays, UV light other forms of high energy radiation – chemical – nucleotide analogs (mimic a NT but incorrectly pair), intercalating agents, others alter chemical properties of the base (alters their pairing capacity) • types of mutations: lecture on transcription and translation – small scale – point mutations – large scale – effect large areas of chromosome sequence

Genes of Cancer • two broad category of genes affected by these: tumorsuppressor genes and proto-oncogenes • tumor suppressor genes – meant to suppress the unrestricted development of cells – repair damaged DNA – control cell adhesion – inhibit the cell cycle using cell-signaling pathways • proto-oncogenes – genes that play roles in normal embryonic development – turned off as development concludes – also play roles in normal growth and division – mutations transform them into cancer causing oncogenes

Mutations in Cancer • genetic mutations can result in the transformation of the genes of cell growth and differentiation • numerous kinds of genetic mutations possible – small scale point mutations, deletions and insertions – large scale deletion or insertions of chromosomal sections, translocation of chromosome sections • transformation of a cancer cell does not always result in increased proliferation – can result in loss of cell-cell adherence + increased ability to invade surrounding tissues – metastatic capacity

Oncogenes and the Cell Cycle • mutation in a proto-oncogene transforms them into an oncogene • oncogenes have abnormal cell cycle control checkpoints • e. g. Cyclin D – in normal cells – cyclin D is a link between G 1 progression – mutations in genes that control cyclin D transcription can result in its overproduction – overproduction of cyclin D in cancer cells drives the cell into and through this phase – even takes cells out of the G 0 phase sooner then they should be – overproduction can result from chromosome translocation - the cyclin D gene is moved next to a very active promoter
- Slides: 51