KANSERN MOLEKLER TEMEL ONKOGENLER Yrd Do Dr Mustafa






 Parental çift heliksin")


, DNA’da saklanan genetik bilgilerin bir RNA molekülü (m.")




, gen ifadesinin son aşamasıdır. Transkripsiyonla RNA’ya kopyalanan genetik")

, GTP varlığında, ribozom alt")
 sayesinde,")












, hematopoietik hücrelerin, inflamatuvar ve")



 belirtileri olmaksızın, bir dizi biyokimyasal değişiklik ile")
 farklı hücre ölümüdür. Oksidan stres, mitokondriden sitokrom")
- Slides: 37
KANSERİN MOLEKÜLER TEMELİ- ONKOGENLER Yrd. Doç. Dr. Mustafa ALTINIŞIK ADÜ Tıp Fakültesi Biyokimya Anabilim Dalı
Kanser, hücre büyümesi ve bölünmesi yani hücre döngüsünü düzenleyen mekanizmalardaki bozukluk sonucu ortaya çıkan patolojik bir durumdur.
HÜCRE DÖNGÜSÜ 1 Hücre döngüsünün dışında, son farklılaşmasını tamamlamış ve bölünmesi duran hücrelerin dinlenme evresi olarak G 0 evresi de düşünülmektedir.
HÜCRE DÖNGÜSÜ 2 S evresi, DNA replikasyonu, kromozomun çiftlenmesi, RNA ve protein sentezinin olduğu evredir. Bu evre 6 -8 saat sürer.
DNA replikasyonu 1 DNA’nın replikasyonu, DNA moleküllerinin doğru kopyalarının yapılmasıdır. Bugün kabul gören görüşe göre DNA’nın replikasyonu semikonservatifdir; bir DNA molekülünün iki kolundan her biri yeni bir DNA kolu sentezi için bir kalıp olarak görev görür ve sonuçta meydana gelen iki yeni DNA molekülü yeni ve eski kollar içerirler
DNA replikasyonu 2 DNA replikasyonu, orijin diye adlandırılan bir başlama noktasında başlar ve genellikle iki yöndeki replikasyon çatallarında 5 3 yönünde ilerler; kalıp olarak görev gören kol 3 5 yönünde okunur
DNA replikasyonu 3 E. coli’de DNA replikasyonu beş aşamada gerçekleşir: 1) Parental çift heliksin açılması. 2) Bir oligonükleotid primerinin sentezi. 3) DNA zincirinin 5 3 yönünde büyümesi. 4) Primerin çıkarılması. 5) Yeni sentez edilen DNA zincirinin birleşmesi
DNA replikasyonu 4 DNA replikasyonunda DNA polimerazların etkisi, kalıp kolun karşısında büyüyen DNA koluna uygun deoksinükleozid trifosfatlardan (d. NTP) deoksinükleozid monofosfatların (d. NMP) girişini sağlamaktır.
DNA replikasyonu 5 Yeni bir DNA şeridinin sentezi tamamlandıktan sonra DNA giraz, replike olmuş DNA’nın tekrar doğal haline kıvrılmasına yardımcı olur Ökaryotik hücrelerde de DNA replikasyonu semikonservatifdir ve çift yönlü cereyan eder; öncü şeritte devamlı, geciken şeritte kesintili olur. Ancak ökaryotik hücrede DNA replikasyonu, kromatin üzerinde binlerce yerde birden başlar ve devam eder
RNA sentezi 1 RNA sentezi (transkripsiyon), DNA’da saklanan genetik bilgilerin bir RNA molekülü (m. RNA, t. RNA, r. RNA) şeklinde kopyalanması veya yazılması olayıdır. Bir RNA molekülü, DNA’nın kalıp kolunun dizilişini bütünleyici ribonükleotidlerin ATP, GTP, CTP ve UTP’tan pirofosfatlar ayrılması suretiyle polimerizasyonu sonucunda, 5 3 yönünde sentezlenir.
RNA sentezi 2 RNA sentezi için RNA polimeraza, başlama ve sonlanma sinyallerine gereksinim vardır
RNA sentezi 3 RNA polimeraz kalıp kol tarafından yönetilen ve baz eşleşmesi kuralları tarafından yorumlanan spesifik bir diziliş içinde ribonükleotidleri polimerize ederken pirofosfatlar serbest bırakılır ve böylece RNA sentezlenir
RNA sentezi 4 Transkripsiyon sonunda oluşan RNA’lar primer RNA’lar diye adlandırılırlar ve genellikle hemen kullanılmazlar; RNA processing diye tanımlanan bazı işlemlerden geçtikten sonra işlev görebilecek olgun RNA’lar haline gelirler
RNA sentezi 5
Protein sentezi 1 Protein sentezi (translasyon), gen ifadesinin son aşamasıdır. Transkripsiyonla RNA’ya kopyalanan genetik bilgi son olarak bir protein veya polipeptit zinciri haline dönüştürülür
Protein sentezi 2 Protein sentezinin üç komponenti m. RNA, t. RNA ve ribozomlardır. m. RNA, proteinin amino asit sırasını belirleyen kodu (şifre) içerir Protein sentezi başlayacağı zaman, sitoplazmada bulunan amino asitler, kendilerine özgü ve Mg 2+ gerektiren aminoaçilt. RNA sentetaz enzimleri yardımıyla kendilerine özgü t. RNA’lara bağlanarak aminoaçil-t. RNA şeklinde aktiflenirler.
Protein sentezi 3 Prokaryotlarda protein sentezi başlarken başlama faktörleri (IF), GTP varlığında, ribozom alt üniteleri, m. RNA ve f. Mett. RNA’dan, m. RNA’nın 5′ ucuna yakın bir bölgesinde başlama kompleksi oluşur
Protein sentezi 4 Başlama kompleksi oluştuktan sonra, GTP’ın hidrolizi ve elongasyon faktörü (EF-Tu) sayesinde, bu kompleksteki A yerine, m. RNA’nın buraya rast gelen kodonunu tamamlayan antikodonu içeren aminoaçil-t. RNA gelir
Protein sentezi 5 ribozomda bulunan peptidil transferaz enziminin katalitik etkisiyle P yerindeki fmett. RNA’da bulunan aminoaçil grubu, A yerindeki aminoaçilt. RNA’nın aminoaçilinin serbest amino grubuna peptit bağı ile bağlanmak üzere taşınır
Protein sentezi 6 GTP’ın hidrolizi ve EF-G (Prokaryotlarda EF-G’nin karşılığı, ökaryotlarda e. EF 2’dir. ) sayesinde P yerindeki t. RNA kompleksten ayrılır, A yerindeki dipeptidil-t. RNA A yerinden P yerine yer değiştirirken ribozom, m. RNA üzerinde 3 ucuna doğru bir kodon ilerler ve A yerine uygun aminoaçilt. RNA gelir
Protein sentezi 7 a Son iki basamaktaki olayların tekrarı sonucunda polipeptit zinciri amino-terminal uçtan karboksilterminal uca doğru uzar Polipeptit zincirinin uzaması sonlandırılacağı zaman, A yerine UAG, UAA, UGA sonlandırma kodonlarından biri gelir; buraya terminasyon faktörü (RF) bağlanır ve önce polipeptidil-t. RNA bağı hidroliz olur daha sonra diğer komponentler dissosiye olurlar
Protein sentezi 7 b
Protein sentezi 8 a Translasyon sonunda yeni sentezlenen polipeptit zincir, biyolojik olarak aktif forma dönüşmek için posttranslasyonal modifikasyonlar denen değişikliklere uğrar. Başlıca posttranslasyonal modifikasyonlar şunlardır: 1) Amino-terminal ve karboksil-terminal modifikasyonlar. 2) Sinyal dizisinin çıkarılması. 3) Bazı özel amino asitlerin modifikasyonu. 4) Karbonhidrat yan zincirlerin bağlanması. 5) İzoprenil grupların eklenmesi. 6) Prostetik grupların eklenmesi. 7) Proteolitik işlem. 8) Disülfid çapraz bağlarının oluşması
Protein sentezi 8 b Çoğu lizozomal protein, membran proteini ve hücreden salgılanan proteinler, kendilerinin endoplazmik retikulum lümeni içinde translokasyonu için işaret olan bir aminoterminal sinyal dizisine sahiptirler. Yeni sentez edilen proteinlerin sinyal dizilerinin çıkarılması ve özel sellüler lokalizasyonlarına trasport edilmesi, protein hedeflenmesi olarak tanımlanır. Glikozilasyon, protein hedeflenmesinde anahtar rol oynar
Protein sentezi 8 c
HÜCRE DÖNGÜSÜ 2 G 2 evresi, S ve M evreleri arasındaki evredir. Bu evrede DNA replikasyonu olmaz; RNA ve protein sentezi devam eder; evre 3 -4 saat sürer ve hücre büyüklüğü iki katına ulaşır. M evresi, mitozun gerçekleştiği evredir. G 1 evresi, ilk bölünmede oluşan eş hücrelerin tekrar hücre bölünmesine girmeden önceki evresidir. Bu evrede DNA replikasyonu olmaz; RNA ve protein sentezi devam eder; evre 3 -4 saat sürer ve hücre S evresine hazırlanır.
HÜCRE DÖNGÜSÜ 3 Hücre döngüsünün dışında, son farklılaşmasını tamamlamış ve bölünmesi duran hücrelerin dinlenme evresi olarak G 0 evresi de düşünülmektedir. G 0 dinlenme evresi, birkaç saat, birkaç gün veya ömür boyu sürebilmekte ve bu evrede hücrede tüm biyokimyasal olaylar aktif bir şekilde devam etmektedir. G 0 dinlenme evresindeki hücre bölüneceği zaman, G 1 evresinin başlangıcından itibaren hücre bölünme döngüsüne girer. Büyüme faktörleri, sitokinler ve tümör virüsleri gibi mitojenik iletiler, G 0 evresindeki hücrenin G 1 evresine girmesine yol açmaktadır.
HÜCRE DÖNGÜSÜ 4 a Hücre büyümesi ve bölünmesi yani hücre döngüsü evrelerini, büyüme faktörleri, sitokinler, onkogenler, siklinler ve sikline bağlı kinazlar (CDK) gibi proteinler koordinasyon içinde düzenlerler. Büyüme faktörleri ve sitokinler, mitojen etkileriyle hücre döngüsünü başlatırlar. Hücre döngüsünün sonlandırılmasını etkileyen faktörlerden biri, hücrenin diğer hücrelerle temas etmesini sağlayan kaderinler ve ekstrasellüler matrikse bağlanmasını sağlayan integrinlerdir.
HÜCRE DÖNGÜSÜ 4 b Siklin-CDK kompleksleri, gerekli proteinleri aktifleyerek hücrenin metabolik aktivitelerini ve hücre bölünmesini düzenlerler. Siklin A, siklin B, siklin C, siklin D gibi en az on farklı siklin ve CDK 1, CDK 2, CDK 3, CDK 4, CDK 5, CDK 6 gibi en az sekiz CDK belirlenmiştir. Siklin-CDK komplekslerinin aktivitelerinin düzenlenmesinde de p 15, p 16, p 21, p 27 gibi CDK inhibitörü proteinler etkili olmaktadır. Bu proteinler, tümör baskılayıcı genler olarak bilinen genlerin ürünleridirler.
HÜCRE DÖNGÜSÜ 4 c Büyüme faktörleri, hücre bölünmesini uyaran veya inhibe eden veya özelleşmiş hücreye farklılaşmasını başlatan peptitlerdir. Epidermal büyüme faktörü (EGF), insülin benzeri büyüme faktörleri (IGF-I ve IGF-II), hematopoietik büyüme faktörü, hücre proliferasyonunda etkilidir. Fibroblast büyüme faktörü (FGF), hücrelerin göç etmelerinde etkilidir. Trombositten türeyen büyüme faktörü (PDGF) ve transforme edici büyüme faktörü (TGFα), yara iyileşmesinde etkilidir.
HÜCRE DÖNGÜSÜ 4 d Sitokinler (hücresel kaynaklı ve çözünebilen moleküller), hematopoietik hücrelerin, inflamatuvar ve immün yanıtın gelişimi ve düzenlenmesinde aracılık eden peptit veya glikoprotein yapıda kimyasal ileti molekülleridir. Sitokinler, genel özellikleri, işlevleri, aile grupları ve primer fonksiyonlarına göre sınıflandırılmaktadır. Aile gruplarına göre sitokinler beş sınıftır: 1) İnterlökinler: IL-1α, IL-1 , IL-2, IL-3, IL-4, IL-5, IL-14, IL-15 2) Kemokinler: IL-18, MCP-1 3) İnterferonlar: IFNα, IFN - IFNγ 4) Sitotoksik/immünodüzenleyici/büyüme faktörleri: TNFα, TNF , TGF 5) Koloni uyarıcı faktörler (hematopoietik büyüme faktörleri): GCSF, GM-CSF, IL-3, IL-7
PROONKOGENLER VE ONKOGENLER Hücrelerin büyüme, çoğalma, farklılaşma ve programlanmış hücre ölümü olan apoptoz için aldıkları iletileri hücre membranından başlamak üzere çekirdeğe kadar sürdürdükleri sinyal ileti mekanizmasında işlev gören birçok proteinin ekspresyonundan sorumlu olan genler, protoonkogenler olarak tanımlanırlar. Normal büyüme ve farklılaşmasını destekleyen genler olan protoonkogenler herhangi bir nedenle mutasyona uğrarlarsa onkogenlere dönüşürler.
ONKOGENLER VE KANSER OLUŞUMU 1 Onkogenler, kanser oluşturan genlerdir. Bilinen yaklaşık 100 kadar onkogen bulunmaktadır. Normalde hücre bölünmesini baskılayan proteinleri (p 15, p 16, p 21, p 27 gibi CDK inhibitörü proteinler) kodlayan ve tümör baskılayıcı genler olarak tanımlanan genlerin (antionkojenler) birinde veya birkaçındaki mutasyon da tümör oluşumuna neden olmaktadır. Apoptozu düzenleyen genlerin mutasyonu da neoplastik hücre birikimine yani kansere yol açmaktadır.
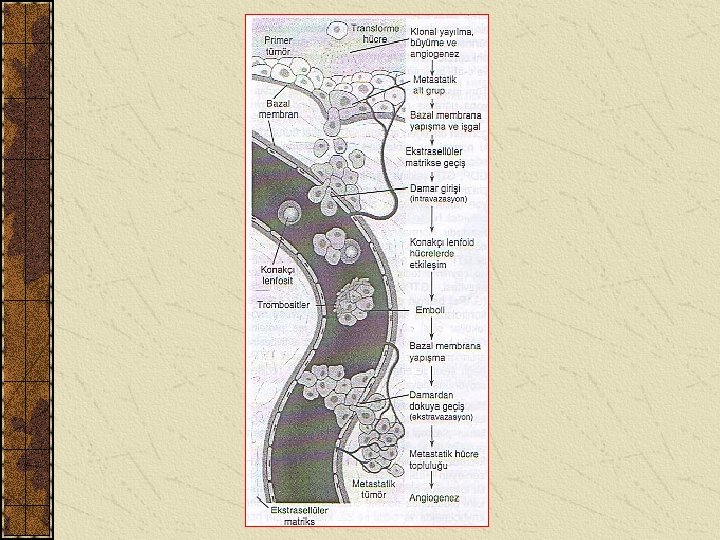
ONKOGENLER VE KANSER OLUŞUMU 2 Normal bir hücrenin kanser hücresi haline dönüşmesine transformasyon denir. Kanser hücrelerinin ulaşacağı son aşama metastatik fenotipdir.
APOPTOZ 1 Apoptoz, pasif hücre ölümü (nekroz) belirtileri olmaksızın, bir dizi biyokimyasal değişiklik ile gözlenen programlanmış hücre ölümüdür. Apoptoz, RNA, protein sentezi ve yeni enzim aktivitelerini gerektiren aktif bir olaydır; pek çok fizyolojik olay ve hastalıkta rol oynar.
APOPTOZ 2 Apoptoz, nekrozdan (pasif hücre ölümü) farklı hücre ölümüdür. Oksidan stres, mitokondriden sitokrom c salınması, sistein proteazların (kaspazlar) aktivasyonu, endonükleazların aktivasyonu gibi moleküler olaylar ve eksternal uyaranlar apoptozu başlatabilmektedir. Hücrede apoptozun düzenlenmesinde sistein proteazlar (kaspazlar) ve Bcl-2 gen ailesi olmak üzere iki protein ailesinin önemli bir rol oynadığı gösterilmiştir.