Interaksi Tanaman dan Bakteri Patogen Oleh Irda Safni

Interaksi Tanaman dan Bakteri Patogen Oleh : Irda Safni

Diseases ? Plant diseases are the result of infection by any living organisms that adversely affect the growth, development, physiological functioning and productivity of a plant, manifesting outwardly as visible symptoms. Any Parasite organisms that cause disease are called pathogens and pathogens on parasitoid is known as host.

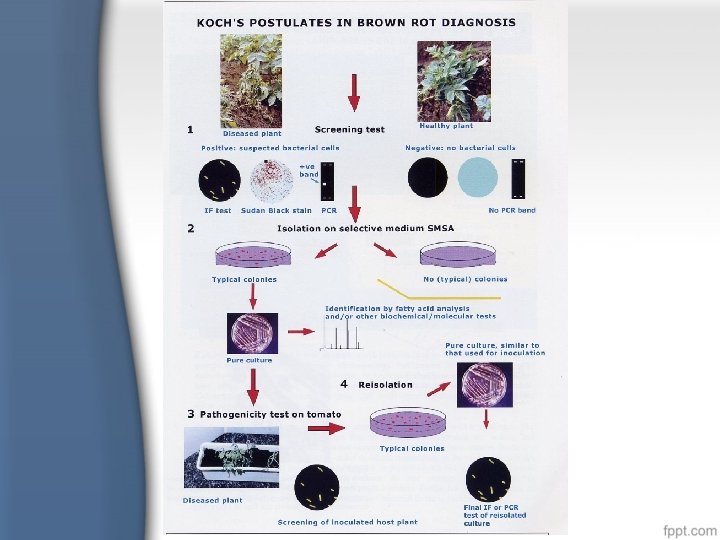
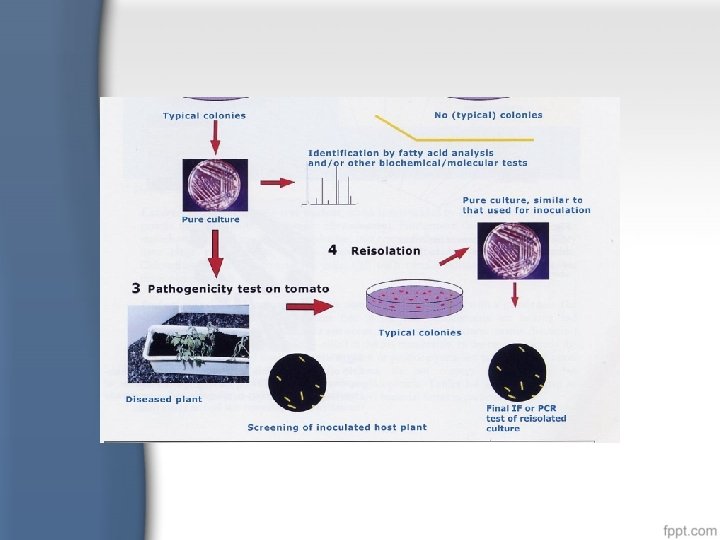
Postulat Koch 1. Mikroba penyebab penyakit harus ada pada individu yang sakit dan tidak terdapat pada individu yang sehat 2. Mikroba penyebab penyakit tsb, harus dapat diisolasi/dibiakkan secara murni 3. Mikroba dari biakan murni tsb, bila diinfeksikan kembali pada individu yang sehat, akan menumbulkan gejala penyakit yang sama 4. Mikroba yang telah diinfeksikan kembali tsb, dapat diisolasikan kembali dan akan mempunyai bentuk yang sama seperti asalnya

Plant – Pathogen Interaction Plants exist in a world filled with bacteria, nematodes, and possibly parasitic plants. fungi, They may be inoculated with viruses during feeding by insects or by other vectors (insects/pest, water, air, humans etc). Plant pathogens have made many adaptations to enable them to invade plants, overcome plant defense mechanisms, and colonize plant tissues for growth, survival, and reproduction.

Plant – Pathogen Interaction Pathogens accomplish these activities mostly through secretions of chemical substances that affect certain components or metabolic mechanisms of their hosts. Penetration and invasion, however, seem to be aided by or in some cases be entirely the result of, the mechanical force exerted by certain pathogens on the cell walls of the plant.

Harold Henry Flor, 1900– 1991 Flor received a Ph. D. degree in 1929 from the University of Minnesota. He worked for the USDA for three years at Washington State University and then for the remainder of his career at North Dakota State University § In the 1940 s, Flor developed the gene-for-gene concept to explain the genetic interactions between Melampsora lini and flax. His theories were put to use in Flor’s own flax breeding program to successfully develop rust-resistant flax. This concept provided the underpinnings for research on the genetics of host-pathogen interactions for the next 70 years.

Gene For Gene Hypothesis § The concept of gene for gene hypothesis was first developed by Flor in 1956 based on his studies of host pathogen interaction in flax, for rust caused by Malampsora lini. § The gene for gene hypothesis states that for each gene controlling resistance in the host, there is corresponding gene controlling pathogenicity in the pathogen. The resistance of host is governed by dominant genes and virulence of pathogen by recessive genes. The genotype of host and pathogen determine the disease reaction. § When genes in host and pathogen match for all loci, then only the host will show susceptible reaction. If some gene loci remain unmatched, the host will show resistant reaction. The gene for gene hypothesis is also known as “Flor Hypothesis. ” § At molecular level, it is considered that gene for gene resistance usually involves production of toxins antibiotic proteins by a resistance gene. Theproduction of toxins is related to gene dosage. § The resistance controlled by domain gene is the most desirable. Gene for gene relationship are rare or unknown for disease caused by viruses, bacteria, Fusarium.


Biochemistry of Gene-for-Gene Hypothesis The Receptor-Elicitor Model of gene-for-gene interactions. The resistance allele of the plant encodes a receptor that recognizes an elicitor produced by the pathogen. Recognition of the pathogen elicitor by the plant receptor initiates plant defense responses that lead to plant resistance. If the pathogen produces the elicitor, it is avirulent. If the pathogen does not produce the elicitor, it is virulent

Mode of Bacterial Infection

Chemical Weapons of Pathogens Some pathogens in plant are largely chemical in nature. Therefore, the effects caused by pathogens on plants are almost entirely the result of biochemical reactions taking place between substances (Hydrolytic enzymes) secreted by the pathogen and those present in or produced by the plant. The main groups of substances secreted by pathogens in plants that seem to be involved in production of disease either directly or indirectly, are enzymes, toxins, growth regulators and polysaccharides (plugging substances).

Chemical Weapons of Pathogens Toxins seem to act directly on protoplast components and interfere with the permeability of its membrane and with its functions. Growth regulators exert a hormonal effect on the cells and their increase or decrease their ability to divide and enlarge. Polysaccharides seem to play a role only in the vascular diseases, in which they interfere passively with the translocation of water in the plants.

Pathogenicity of Bacterial Enzymes That Degrade Cell Walls Plant cell walls are composed of three major polysaccharides: - Cellulose - Hemicellulose - Pectins and, - Lignin, in woody and some other plants. The number of genes encoding cell wall-degrading enzymes varies greatly in the different plant pathogenic bacteria: Softrotting Erwinias produce a wider range of enzymes able to degrade plant cell wall components than any other plant pathogenic bacteria.
 Pectinases: Pectin substances constitute the main components of the middle lamella i.")
ENZYMES (A) Pectinases: Pectin substances constitute the main components of the middle lamella i. e. the intercellular cement that holds in place the cells of plant tissues. Several enzymes degrade pectic substances and are known as pectinases or pectolylic enzymes. Pectinases are believed to be the most important in pathogenesis, as they are responsible for tissue maceration by degrading the pectic substances in the middle lamella and, indirectly, for cell death.

1. Pectic enzymes is pectin methyl-esterases, which removes small branches off the pectin chains. 2. Pectic enzymes is a chain splitting pectinases called polygalacturonases. It split the pectic chain by adding a molecule of water and breaking the linkage between two galacturonan molecules. 3. Pectin lyases is split the chain by removing a molecule of water from the linkage, there by breaking it and releasing products with an unsaturated double bond. Examples of pathogens include Ralstonia solanacearum.
 Cellulases: Ø Cellulose is also a polysaccharide, but it consists of chains of")
(B) Cellulases: Ø Cellulose is also a polysaccharide, but it consists of chains of glucose (1 -4) β-D-glucan molecules. Several saprophytic bacteria cause the breakdown of most of the cellulose decomposed in nature. ØIn living plant tissues, however, cellulolytic enzymes secreted by pathogens play a role in the softening and disintegration of cell wall material.

MICROBIAL TOXINS ØToxins are metabolites that are produced by invading microorganisms and act directly on living host protoplast, seriously damaging or killing the cells of the plant. ØSome toxins act as a general protoplasmic poisons and affect many species of plant representing different families. Others are toxic to only a few plant species or varieties and are completely harmless to others. Many toxins exist in multiple forms that have different potency. (A) Non-host specific toxin (B) Host specific toxin

Bacterial Toxins as Pathogenicity Factors Toxins have been known for a long time to play a central role in parasitism and pathogenesis of plants by several plant pathogenic bacteria. Pseudomonas syringae, P. syringae pv. tomato, and P. syringae pv. maculicola are primarily associated with production of the phytotoxin coronatine. Coronatine functions primarily by suppressing the induction of defense-related genes, but, as happens with most bacterial phytotoxins, it does not seem to be essential for pathogenicity by all strains. The bacterium P. syringae, along with its pathovars, produces several pathotoxins, including syringomycin. Albicidins, produced by Xanthomonas albilineans, block the replication of prokaryotic DNA and the development of plastids, thereby causing chlorosis in emerging leaves. Albicidins interfere with host defense mechanisms and thereby the bacteria gain systemic invasion of the host plant.

Extracellular Polysaccharides as Pathogenicity Factors Ø Fungi, bacteria, nematodes and possibly other pathogens constantly release varying amounts of mucilaginous substances that coat their bodies and provide the interface between the outer surface of the microorganism and its environment. Ø The role of the slimy polysaccharides in plant disease appears to be particularly important in wilt diseases caused by pathogens that invade the vascular system of the plant. Ø Large polysaccharide molecules released by the pathogen in the xylem may be sufficient to cause a mechanical lockage of vascular bundles and thus initiate wilting.
 play an important role in pathogenesis of many bacteria by both")
Extracellular polysaccharides (EPS) play an important role in pathogenesis of many bacteria by both direct intervention with host cells and by providing resistance to oxidative stress. In the bacterial wilt of solanaceous crops caused by Ralstonia solanacearum, EPS 1 is the main virulence factor of the disease. EPS 1 is produced by the bacterium in massive amounts and makes up more than 90% of the total polysaccharide. EPS likely causes wilt by occluding the xylem vessels and by causing them to rupture from the high osmotic pressure.

Growth Regulators Ethylene – • In the fruit of banana infected with Ralstonia solanacearum, the ethylene content increases proportionately with the (premature) yellowing of the fruits, whereas no ethylene can be detected in the healthy fruits.

Polysaccharides ØFungi, bacteria, nematodes and possibly other pathogens constantly release varying amounts of mucilaginous substances that coat their bodies and provide the interface between the outer surface of the microorganism and its environment. ØThe role of the slimy polysaccharides in plant disease appears to be particularly important in wilt diseases caused by pathogens that invade the vascular system of the plant. ØLarge polysaccharide molecules released by the pathogen in the xylem may be sufficient to cause a mechanical lockage of vascular bundles and thus initiate wilting.

PATOGENESIS BAKTERI PATOGEN TUMBUHAN Tahap-tahap Infeksi I. MIGRASI Definisi : Berpindahnya sel/ massa sel bakteri ke arah inang atau bagian inang yang sesuai Faktor yang berperan a. Bakteri Flagel yg berkorelasi dengan motility (kemampuan untuk bergerak. Flagel untuk membantu mendayung saat bergerak Gen pembentukan flagella pd bakteri bersifat on / off (bisa terbentuk atau tidak) Perannya penting jika bakteri belum masuk jaringan inang, jika sudah masuk flagel tidak dibentuk Flagel bisa digunakan bila ada filum air

b. Lingkungan Filum air bebas, umumnya penyakit bakteri akan timbul jika kelembapan tinggi c. Faktor Inang - eksudat akar/ sekresi ; pintu buangan sisa metabolisme yang tidak dipakai oleh inang tetapi diperlukan oleh bakteri sebagai sumber nutrisi - Bakteri dapat mengeliat / mendekat ke eksudat akar karena konsentrasi di sekitar eksudat akar lebih tinggi dari konsentrasi bakteri d. Arah migrasi luka pada inang yang dituju bakteri adalah : 1. Luka alami (luka pertumb tanaman : tunas, cabang akar), buatan (luka alat pertanian, cuaca, serangga 2. Lubang alami (stomata, hidatoda, lenti sel, nectar)
 Definisi : Proses kontak / pengenalan sel bakteri dengan jaringan inang")
II. PENGENALAN (RECOGNITION) Definisi : Proses kontak / pengenalan sel bakteri dengan jaringan inang sebelum infeksi terjadi. Pada saat kontak maka terjadi hubungan yang spesifik antara reseptor pada sel inang dan molekul pada permukaan sel bakteri Syarat : Harus ada kompatibelitas antara dinding sel inang dan bakteri sering terjadi karena substansi yang kompatibel (misal : Lektin / LPS)
 LPS")
Contoh : 1. Agrobacterium tumefaciens LPS vs receptor sel inang (inang harus luka) LPS Mikrofibril Mikrokoloni (enzim pemecah senyawa pectin) Sell inang rusak TIP = Tumor Inducing Plasmid (DNA plasmid) DNA bakteri masuk Mengacaukan mekanisme DNA Inang Pembelahan sel inang banyak sekali (gall)
 Parenkim plasmolisis Dinding xylem pecah Xylem")
2. Ralstonia solanacearum EPS atau enzim EG (Endoglukonase) Parenkim plasmolisis Dinding xylem pecah Xylem Menyumbat fungsi xylem - massa bakteri - sisa degradasi sel (makromolekul)

III. PENETRASI 1. Lewat hidatoda / tepi daun misal : penyakit busuk hitam pada kubis Xanthomonas campestris pv. campestris gejala dari tepi lewat hidatoda Xanthomonas oryzae pv. oryzae pada padi gejala diawali dari pinggir daun, bakteri masuk lewat air gutasi 2. Lewat stomata berbanding lurus dengan jumlahnya tetapi juga tergantung struktur stomata - lebar / kecinya celah - tebal / menonjolnya mulut stomata misal : Citrus paradis lebih rentan terhadap X. campestris pv. citri daripada Citrus reticulata (jeruk keprok) karena celah stomata lebih lebar

3. Lewat kelenjar madu Terjadi pada tanaman Apel / Pear Oleh Erwinia amylovora Kelenjar madu yang terbuka lebar lebih cepat terserang Pada Pear melalui kelenjar madu Apel kepala putik 4. Lewat lenti sel Lenti sel : hubungan antara sel yang satu dengan yang lain longga Lapisan filogen selalu membelah sehingga sel epidermis akan terdesak pecah mudah terlewati bakteri Contoh : Streptomyces scabies 5. Lewat luka - alat pertanian -serangga - retakan tempat pada trichoma - retakan tempat keluarnya rambut akar contoh : Ralstonia solanacearum

Bacterial Adhesion to Plant Surfaces Most bacteria do not need adhesion mechanisms except perhaps when they are moving through the xylem and phloem. However, the crown gall bacterium Agrobacterium, requires attachment to plant surface receptors as the first step in the transfer of T-DNA and development of disease symptoms. Several other plant pathogenic bacteria also have genes that encode proteins likely to be involved in attachment and aggregation. Xylella, Xanthomonas, and Ralstonia, all colonizing plant vessels at some stage of infection, also contain additional adhesin gene homologs and homologs of hemagglutinin-related genes found in many bacteria pathogenic to mammals.

Bacterial Secretion Systems Secretion systems are essential pathogenicity tools for bacteria because they make possible the translocation of bacterial proteins and other molecules into host plant cells. There are five forms of secretion systems on the basis of the proteins that form them, i. e. 1. Type I-SS Is present in almost all plant pathogenic bacteria and carries out the secretion of toxins such as hemolysins, cyclolysin, and rhizobiocin. They consist of ATP-binding cassette (ABC) proteins and are involved in the export and import of a variety of compounds through energy provided by the hydrolysis of ATP.

2. Type II-SS is common in gram-negative bacteria and is involved in the export of various proteins, enzymes, toxins, and virulence factors. 3. Type III-SS is the most important in terms of pathogenicity of the bacteria in the genera Pseudomonas, Xanthomonas, and Ralstonia. The primary function of type III-SS is the transport of effector proteins across the bacterial membrane and into the plant cell. 4. Type IV-SS transports macromolecules from the bacterium to the host cell. The proteins transferred are very similar to those responsible for the mobilization of plasmids among bacteria. 5. The type V-SS autotransporter is found in Xylella and Xanthomonas and contains genes that encode surfaceassociated adhesins.

The contact between plant and pathogenic microorganism lead to a particular chain of events in the plant organism. Communication requires a mobile ‘systemic signal’. Interaction lead to two type of response: 1) Compatible response 2) Incompatible response
 Host Reaction")
Patogenesis & Respon Tanaman Pathogens Pre-penetration and Penetration Invasion Colonization Diseases (Symptoms) Host Reaction Naturally occurring Pre-existing defence structure Biochemical defence Post infectional Structural defence Biochemical defence

Plant defense constitutive • • Cell walls Waxy epidermal cuticle Bark Thorns, Spine Inducible • • Toxic chemicals Pathogen degrading enzymes Deliberate cell suicide Systemic resistance
 Induced structural barriers Induced defence phytoalexins Pathogenesis related")
Induced defence Programmed cell death (PCD) Induced structural barriers Induced defence phytoalexins Pathogenesis related proteins (PR-proteins) Post transcriptional gene silencing (PTGS) Heil, M. , and R. M. Bostock. 2002

Hypersensitive response HR lesions Rapid, localized plant cell death upon contact with avirulent pathogens. HR is considered to be a key component of multifaceted plant defense responses to restrict attempted infection by avirulent pathogens. Host cell recognizes the bacterium and initiates programmed cell death to restrict the growth of the pathogen, which thus does not cause disease. HR also contributes to the establishment of the long-lasting systemic acquired resistance against subsequent attack by a broad range of normally virulent pathogens.
 • Disruption of cell")
HR Includes: • Oxidative bust (production of reactive oxygen species) • Disruption of cell membranes • opening of ion channels • Cross linking of phenolics with cell wall component • Production of anti-microbial phytoalexins and PR protein • apoptosis (programmed cell death)

Programmed Cell Death • Programmed cell death is a genetically regulated process of cell suicide that is central to the development, homeostasis and integrity of multicellular organisms

Purpose of Cell Death • Cells that are produced in excess • Cell that have no function • Cell that develop improperly • Cell that have stop further infection • Cells that are harmful
 SAR is a mechanism of induced defense that confers long-lasting")
Systemic Acquired Resistance (SAR) SAR is a mechanism of induced defense that confers long-lasting protection against a broad spectrum of microorganisms. Enhance resistance against subsequent attack by a wide array of pathogen. The vasculature provide the excellent channel for transport of systemic signals. SAR induces the chromatin modification.

• Pathogen recognition triggers a number of rapid cellular responses, including ionic changes, and phosphorylation cascades, which precede the accumulation of reactive oxygen species, nitric oxide, and salicylic acid (SA) and the transcriptional activation of defence-related genes. • SA also has a key role in establishing local and systemic resistance to many virulent biotrophic pathogens.

Systemic signals in plant defence These mobile signals help in activation of SAR. Mostly metabolites function as systemic signal, contributing to long distance signaling in plant defence.
 Pathogenesis related proteins, called PR- proteins- A group of plant coded")
Pathogenesis-Related Proteins (PRproteins) Pathogenesis related proteins, called PR- proteins- A group of plant coded proteins Are structurally diverse group toxic to invading pathogens. Produced under stress They are widely distributed in plants in trace amounts but are produced in high concentration following pathogen attack

Significance of PR-proteins • They show strong antifungal and other antimicrobial activity. • These are associated with strengthening of the host cell wall and its out growths and papillae. • Some of the PR-proteins, for example, B-1, 3 glucanase and chitinase, diffuse towards and affect (break down) the chitin-supported structure of the cell walls of several plant pathogenic fungi, whereas lysozymes degrade the glucosmine and muramic acid components of bacterial cell walls.

TERIMA KASIH
- Slides: 49