Il sistema visivo La maggior parte degli oggetti




 dell’occhio Acquoso, serve come nutrimento all’iride e al cristallino")

 rispetto all’emergenza del nervo ottico")

 Diametro antero-posteriore o sagittale (mm) 24, 124, 2 Diametro")




















,")










 pigmentato: ha una duplice funzione. Impedisce una riflessione della luce sulla")


 e i bastoncelli (120 milioni) sono i trasduttori della")










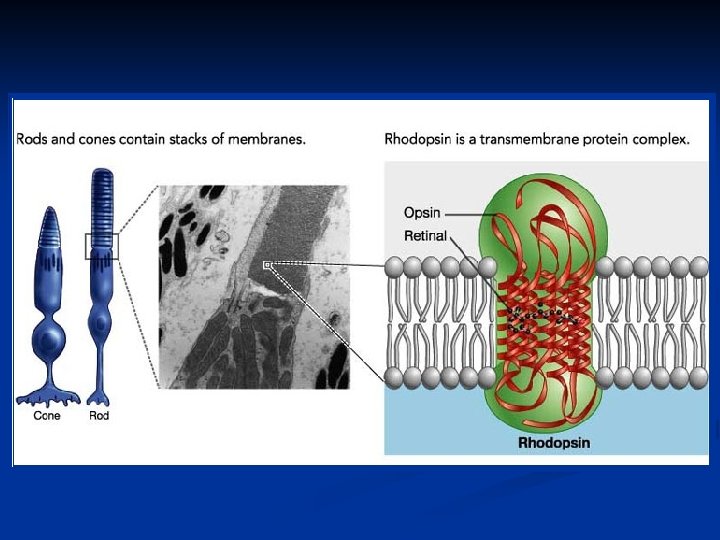





















































. Ogni nervo ottico è")













- Slides: 144
Il sistema visivo
La maggior parte degli oggetti riflette la luce e poiché la luce si propaga ad elevata velocità, è possibile in maniera praticamente istantanea, stabilire dimensione, posizione nello spazio, velocità e direzione del movimento di un oggetto. I raggi luminosi che provengono da un oggetto sono raggruppati e focalizzati su una matrice di fotorecettori. Le attività indotte nei diversi fotorecettori dalla luce interagiscono per dare una rappresentazione BIDIMENSIONALE dell’oggetto che è trasmessa al cervello. Il cervello successivamente ricostruisce un’immaginee TRIDIMENSIONALE uitlizzando le informazioni provenienti dai due occhi.
I prodotti finali dell’attività del sistema visivo sono sensazioni che rappresentano l’oggetto e ciò che lo circonda. Queste sensazioni sono utilizzate per guidare il nostro comportamento immediato oppure possono essere immagazzinate per referenza futura. Le sensazioni visive contengono una gran quantità di informazioni che non è facile comprendere. Il miglior modo per addentrarsi è cominciare dall’occhio stesso.
Anatomia dell’occhio L’iride è il diaframma dell’occhio, in quanto regola la quantità di luce che entra attraverso la pupilla
Cornea: max potere ditottrico (fisso) dell’occhio Acquoso, serve come nutrimento all’iride e al cristallino Viscoso, conferisce forma all’occhio
Rigida, fibrosa Trasparente (ca 12 mm diametro, spessa 0. 5 mm al centro e da 0. 75 a 1 mm ai bordi)
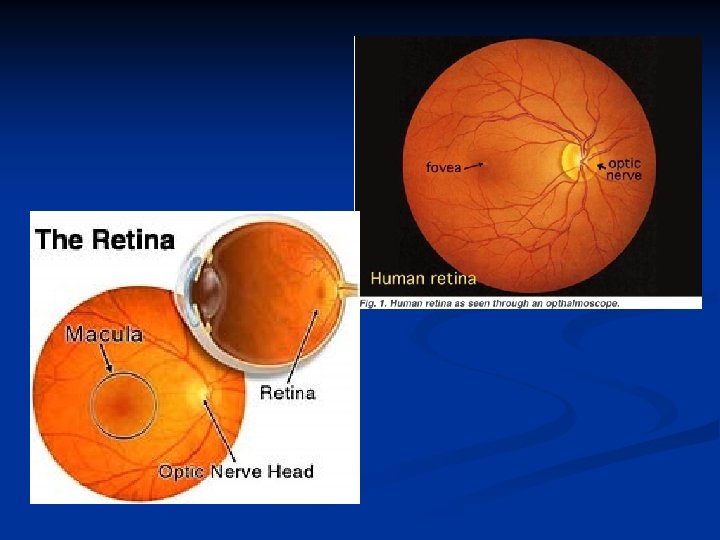
La fovea è in posizione laterale (temporale) rispetto all’emergenza del nervo ottico
La fovea si trova spostata 4 mm lateralmente e 1 mm inferiormente rispetto al polo posteriore.
Occhio: principali misure (valori medi) Diametro antero-posteriore o sagittale (mm) 24, 124, 2 Diametro verticale (mm) 23, 223, 5 Diametro trasversale (mm) 23, 023, 6 Distanza interpupillare (cm) 6 Peso (g) 7 -9 Pressione intraoculare (mm. Hg) 18 -20 Angolo fra asse anatomico ed asse visivo (°) 5 -7 Angolo fra gli assi anatomici dei due occhi (°) 10 Angolo fra asse anatomico ed asse 18
La congiuntiva è lo strato più interno delle palpebre e lo strato più esterno dell’occhio. Si tratta di una membrana mucosa trasparente, molto sottile a funzione protettiva. La cornea è un rivestimento fibroso, relativamente spesso, non vascolarizzato e trasparente. Ha un elevato potere diottrico, ma fisso. È la parte anteriore della sclera e ha la funzione di permettere il passaggio della luce.
La sclera è formata da tessuto connettivo denso e conferisce la forma al globo oculare, rendendolo più rigido. Protegge la parte più interna dell’occhio. Al di sotto della sclera troviamo la coroide. La coroide è riccamente vascolarizzata e fornisce nutrienti alla retina. Ha un aspetto bruno/nero dovuto alla presenza dei melanociti che secernono melanina. E' riccamente vascolarizzata, e serve alla nutrizione dell'epitelio pigmentato e degli strati esterni della retina. Insieme all'epitelio pigmentato della retina, concorre col proprio pigmento ad assorbire i raggi luminosi che hanno attraversato la retina, impedendone la riflessione.
La luce n La luce è una forma di energia radiante emessa dalla materia quando si trova ad alta temperatura, quando è elettricamente eccitata, e quando è sottoposta a certi processi chimici e fisici. Ha caratteristiche fisiche particolari comportandosi come un’onda e anche come un insieme di particelle. Lo spettro luminoso va da 1 nm fino a 104 m.
Ambito dinamico del sistema visivo: la luce visibile
Ottica della visione La luce si propaga in linea retta con una certa velocità a meno che non cambi mezzo di propagazione. Quando cambia mezzo cambia anche la velocità. Questo fenomeno è noto come rifrazione. Si intende come indice di rifrazione (IR) di un mezzo il rapporto fra: velocità della luce nel vuoto / velocità della luce nel mezzo Material IR Air 1. 0003 Water 1. 33 Glycerin 1. 47 Immersion Oil 1. 515 Material IR Glass 1. 52 Zircon 1. 92 Diamond 2. 42 Lead Sulfide 3. 91
Se la luce colpisce un’interfaccia il cui piano è inclinato rispetto alla direzione dei raggi, essa subisce non solo una variazione di velocità , ma anche una deviazione se i due mezzi hanno un differente indice di rifrazione. Il grado di rifrazione aumenta in funzione di: § Rapporto degli indici di rifrazione dei due mezzi § Grado di inclinazione tra il piano dell’interfaccia e fronte d’onda del raggio incidente.
La rifrazione della luce visibile è una caratteristica importante delle lenti, che permette di focalizzare un fascio di luce in un singolo punto. Vediamo gli effetti che una lente convergente, come è il cristallino nell’occhio umano, produce sui raggi luminosi. Consideriamo un fascio di raggi paralleli all'asse principale. Si sperimenta facilmente che tutti i raggi vengono rifratti in modo da convergere in un unico punto detto FUOCO della lente.
Per costruire geometricamente l'immagine prodotta da una lente convergente è sufficiente tenere presente che: § il raggio passante per il centro ottico attraversa la lente senza essere deviato. § un raggio parallelo all'asse principale viene deviato sul fuoco della lente. Il fuoco è la distanza dal centro della lente al punto in cui convergono i raggi provenienti dall’oggetto.
La principale differenza di una lente divergente rispetto ad una convergente è costituita dal fatto che se consideriamo un fascio di raggi paralleli all'asse principale della lente, essi verranno rifratti non in modo da intersecarsi in un punto, ma in modo da "divergere" senza mai intersecarsi. Se però consideriamo i prolungamenti dei raggi rifratti, essi occorrono in un punto sull'asse principale. Dunque il fuoco di una lente divergente è un punto virtuale, nel senso che è determinato non direttamente dai raggi rifratti, ma dai loro prolungamenti.
Per lente positiva si intende una lente convergente ovvero in grado di aumentare la convergenza dei raggi di luce incidenti. Se i raggi di luce incidenti sono paralleli, quelli emergenti convergeranno in un punto (detto fuoco). Per lente negativa si intende una lente divergente ovvero in grado di diminuire la convergenza dei raggi di luce incidenti. Se i raggi di luce incidenti sono paralleli, quelli emergenti divergeranno. Il prolungamento dei raggi emergenti convergono in un punto (il fuoco).
Si definisce diottria il rapporto 1 m / distanza focale ed è una misura del potere di rifrazione di una lente. Tanto più la lente è convessa, tanto maggiore è la deformazione dei raggi. Quindi se il fuoco della lente A è a 1 m, allora la lente avrà il potere di +1 diottria. Se il fuoco della lente B è a 0. 5 m allora la lente B avrà il potere di +2 diottrie. 1 m 0. 5 m +1 diottria A A +2 diottria B
L’occhio è otticamente uguale ad una comune macchina fotografica, in quanto è costituito da un sistema di lenti, da un sistema ad apertura variabile (la pupilla) e dalla retina che corrisponde alla pellicola impressionabile. Il sistema di lenti è costituito da: § Interfaccia aria- superficie anteriore della cornea § Interfaccia superficie posteriore della cornea – umor acqueo § Interfaccia umor acqueo – superficie anteriore del cristallino § Interfaccia superficie posteriore del cristallino – umor vitreo
Se si sommano gli effetti rifrattivi di tutte le superfici rifrangenti dell’occhio considerandole come un’unica lente si ha un potere di 59 diottrie quando il cristallino è accomodato per la visione a distanza. La maggior parte di questo potere è attribuibile alla cornea che ha però un potere rifrattivo fisso. Il cristallino è responsabile per 20 diottrie circa, ma ha un potere di curvatura variabile.
Il potere di rifrazione del cristallino in un soggetto giovane varia fra 20 e 34 diottrie. Questo è possibile grazie alla capacità del cristallino di modificare la sua forma, cioè la sua convessità. In condizioni di riposo il cristallino è sotto tensione e mantenuto appiattito: questa è una condizione atta a mettere a fuoco gli oggetti a distanza
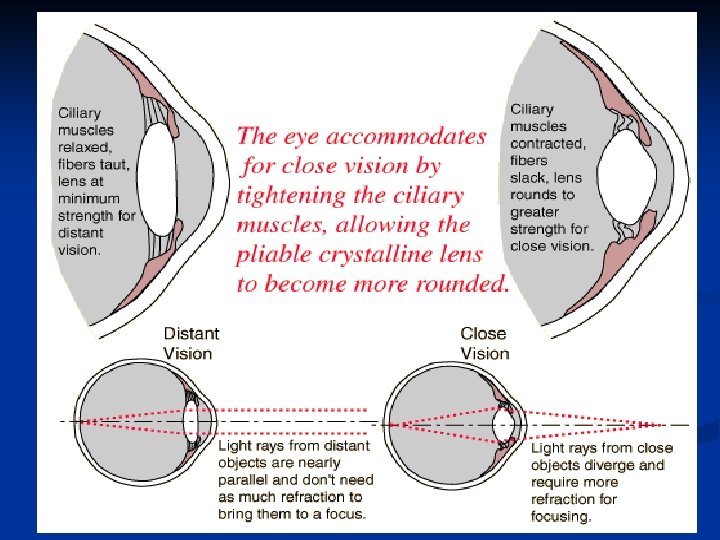
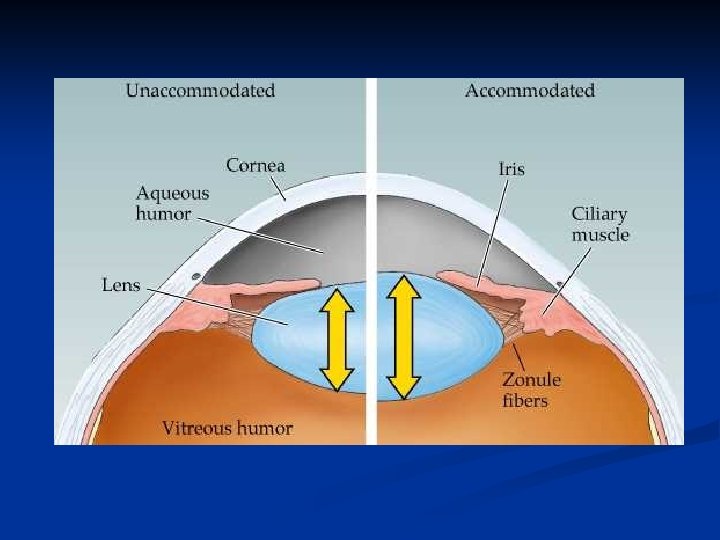
Il cristallino è molto elastico e la sua grande elasticità consente alle fibre che lo tengono sospeso al muscolo ciliare di cambiare la sua forma: questo modifica il suo potere diottrico, rendendo possibile la messa a fuoco a varie distanze. Questo meccanismo è chiamato accomodazione
Il cristallino è formato da una capsula elastica ripiena di materiale viscoso. Nello stato di rilasciamento, quando la capsula non è sottoposta a tensione, il cristallino assume forma sferica. Radialmente intorno al cristallino esistono circa 70 legamenti (legamenti sospensori) che esercitano sui bordi una trazione diretta verso la coroide. Questi legamenti sono costantemente contratti così che il cristallino a riposo assume forma appiattita. Nel punto di inserzione sulla coroide i legamenti sono collegati al muscolo ciliare. Quando esso si contrae i legamenti si detendono e il cristallino si arrotonda.
Non accomodato: muscolo ciliare rilasciato legamenti sospensori tesi cristallino appiattito minimo potere diottrico visione a distanza Accomodato: muscolo ciliare contratto legamenti sospensori rilasciati cristallino arrotondato max potere diottrico visione vicino
Quindi: Muscolo ciliare rilasciato Muscolo ciliare contratto Cristallino appiattito Cristallino sferico Potere diottrico minimo (20 diottrie) massimo (34 diottrie)
Il muscolo ciliare è controllato dal sistema parasimpatico. Quando si ha stimolazione parasimpatica il muscolo ciliare si contrae, i legamenti si detendono e il cristallino diviene più sferico aumentando il suo potere diottrico e mettendo a fuoco gli oggetti più vicini. Normale Accomodato Scarica parasimpatica in aumento all’avvicinarsi dell’oggetto
Il controllo del muscolo ciliare è mantenuto dal SNA attraverso sinapsi colinergiche atropina-sensibili in seguito a stimoli provenienti dalla retina stessa. Presumibilmente le vie implicate in questo meccanismo prevedono: Retina Nervo ottico Nucleo genicolato laterale del talamo Corteccia visiva Collicolo superiore Nucleo oculomotore
Emmetropia: condizione normale in cui, con il cristallino completamente appiattito (muscolo ciliare completamente rilasciato), si mettono a fuoco gli oggetti all’infinito. All’avvicinamento dell’oggetto subentra il meccanismo dell’accomodazione. Occhio emmetrope a riposo Occhio emmetrope accomodato Visione da lontano Visione da vicino
Con il passare degli anni il potere diottrico dell’occhio diminuisce con il risultato che gli oggetti vicini non Età vengono più messi a fuoco. 10 -30 anni Questa condizione pende il nome di presbiopia. Il punto più vicino (near point) che può essere messo a fuoco varia in media in questo modo: Near point 9 cm 45 anni 28 cm 60 anni 80 cm
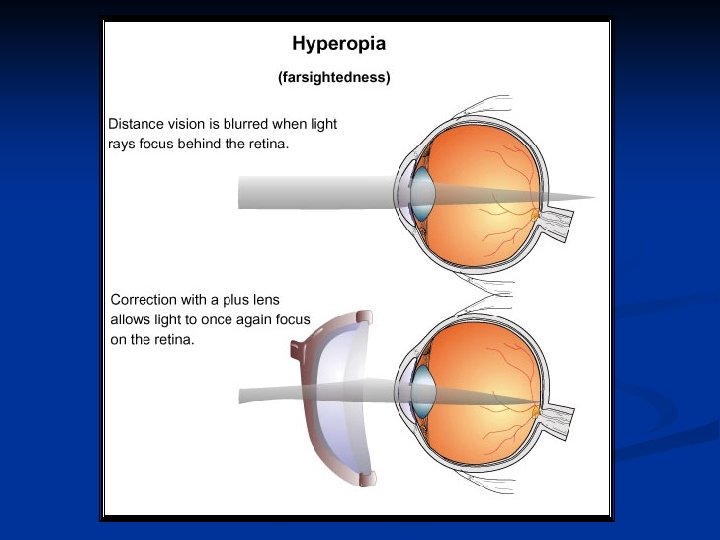
Ipermetropia: il sistema diottrico dell’occhio non riesce a mettere a fuoco gli oggetti lontani senza accomodazione. Quando l’oggetto si avvicina molto il sistema di accomodazione è saturato e non riesce a compensare ulteriormente. Può dipendere sia da una debolezza del sistema diottrico, sia da un asse antero-posteriore dell’occhio troppo corto.
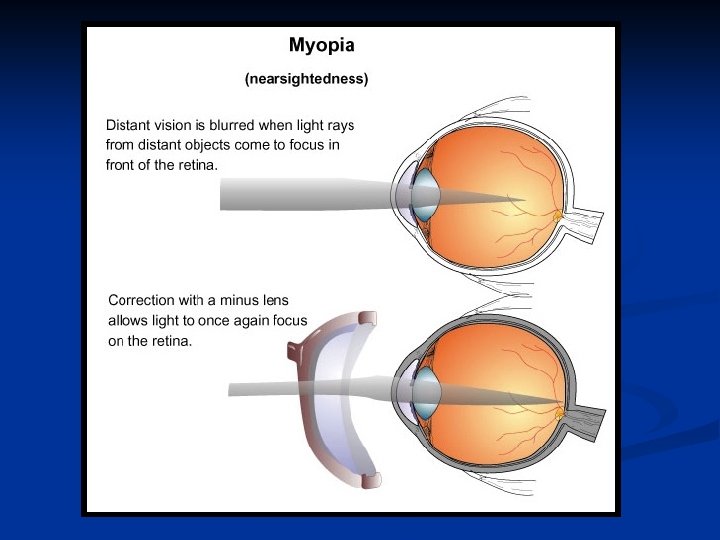
Miopia: in questo caso anche quando il cristallino è completamente appiattito, l’occhio non riesce a mettere a fuoco gli oggetti lontani. Il potere diottrico non può scendere al di sotto del valore minimo, cioè di quando il muscolo ciliare è completamente rilasciato. In genere la miopia dipende da una lunghezza eccessiva dell’asse antero-posteriore Miope Normale dell’occhio.
Astigmatismo: è un difetto di rifrazione dovuto al fatto che il cristallino o più frequentemente la cornea esercitano un potere diottrico differente a seconda del piano considerato. Risultano distorti gli oggetti vicini e quelli lontani. L’accomodazione non funziona perché la curvatura del cristallino varia in ugual misura sui due piani.
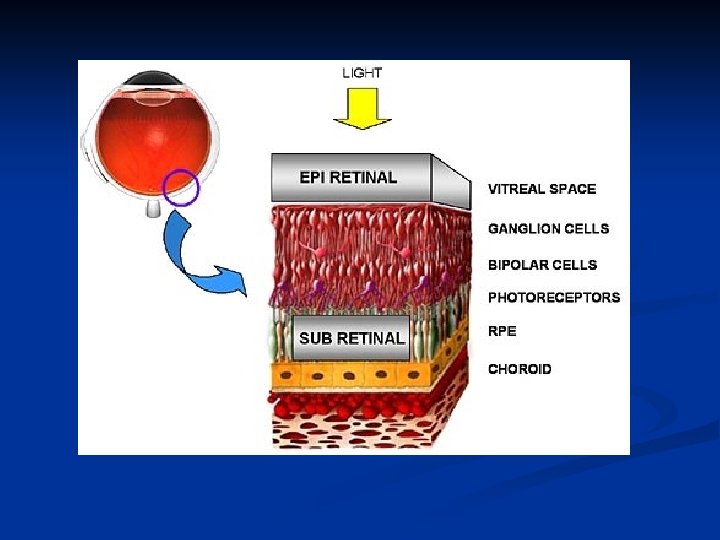
La retina è la più interna delle tre membrane che formano le pareti del globo oculare e si estende dal punto di entrata del nervo ottico fino al margine pupillare dell'iride. Per la sua origine, per la struttura della sua parte principale e per le sue connessioni con il nervo ottico, deve essere considerata come di natura nervosa. È la parte fotosensibile dell’occhio. Contiene i coni responsabili della visione a colori e i bastoncelli responsabili della visione in bianco e nero.
Emisferi cerebrali, con corteccia e parte centrale, gangli della base, ippocampo, corpo striato e sistema olfattivo Talamo, epitalamo, ipotalamo, subtalamo, neuroipofisi, epifisi, retina, nervo ottico, corpi mammillari
I coni e i bastoncelli sono quindi i fotorecettori e, una volta eccitati, questi veicolano l’informazione ad una serie di neuroni che vedremo e da questi al nervo ottico fino alla corteccia cerebrale. La retina è una sottile membrana di colore rosso-arancio dal cui centro si diparte il nervo ottico. A livello del nervo ottico partono i vasi sanguigni principali della retina, l’arteria e la vena centrale della retina, mentre, esattamente al centro della retina, si trova una piccola area di forma ovale, di colore rossastro e priva di vasi sanguigni detta fovea. La parte di retina che comprende la fovea e si estende con un diametro di 3 mm rispetto ad essa è chiamata macula.
La macula ha un colore bruno-giallastro a causa del riflesso della xantofilla, un particolare pigmento contenuto in abbondanza nei coni che ha una funzione di filtro per le radiazioni luminose di bassa lunghezza d’onda. Infatti, poiché la fovea è essenziale per la visione, è importantissimo avere dei meccanismi di difesa per evitare i danni prodotti dalla luce intensa e, soprattutto, dalle radiazioni ultraviolette.
La zona subito attorno alla fovea è chiamata retina centrale mentre la parte ad essa esterna è indicata come retina periferica. In totale la retina è un disco circolare con un diametro di circa 42 mm che finisce in corrispondenza del margine pupillare dell’iride. Vediamo adesso gli strati della retina.
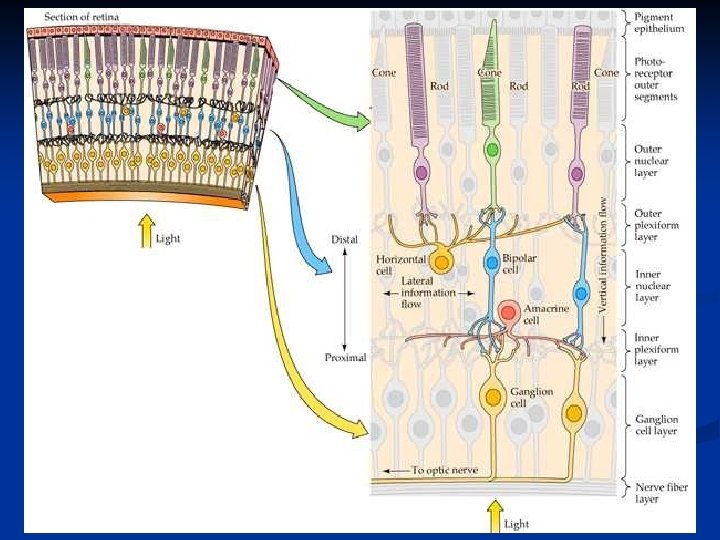
1. Strato (epitelio) pigmentato: ha una duplice funzione. Impedisce una riflessione della luce sulla retina ed è la sede in cui viene riformato continuamente il pigmento visivo. 2. Strato dei fotorecettori: segmenti esterni di coni e bastoncelli 3. Membrana limitante esterna 4. Strato granulare (o nucleare) esterno: corpi cellulari dei fotorecettori 5. Strato plessiforme esterno: sinapsi fra fotorecettori, bipolari e orizzontali
5. Strato granulare interno: corpi cellulari delle bipolari e amacrine 6. Strato plessiforme interno: sinapsi fra bipolari, amacrine e gangliari 7. Strato delle cellule gangliari: corpi somatici delle gangliari 8. Strato delle fibre nervose: formazione del nervo ottico con gli assoni delle gangliari. 9. Membrana limitante interna
Quello che è curioso notare è la direzione in cui la luce attraversa l’occhio. In effetti i fotorecettori sono le ultime cellule a venire colpite dalla luce, perché costituiscono uno strato molto esterno rispetto a quello delle cellule degli altri strati.
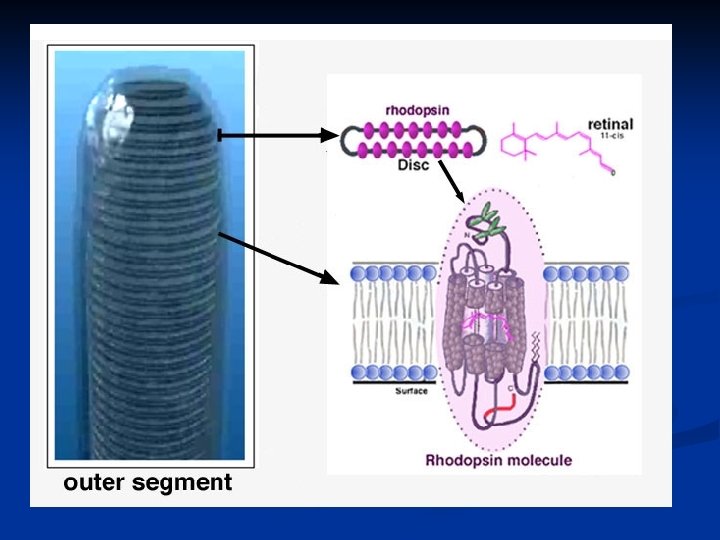
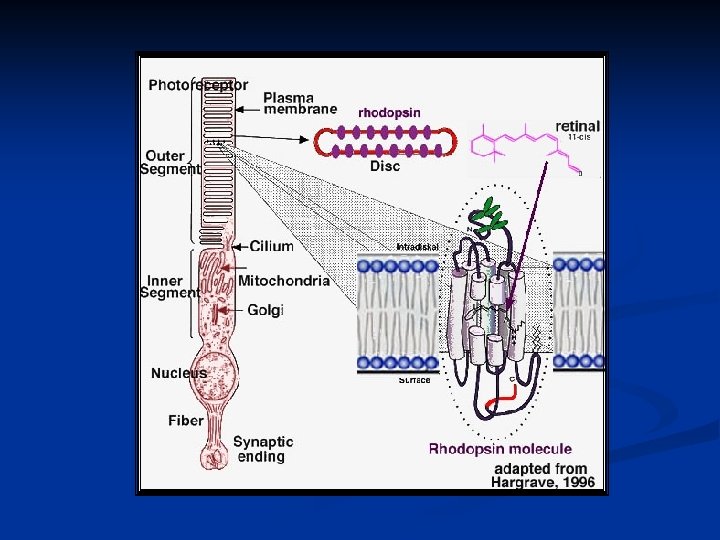
I coni (6 -7 milioni) e i bastoncelli (120 milioni) sono i trasduttori della luce. Sono cellule altamente specializzate caratterizzate da strutture molto particolari per espletare il loro compito. Sono strutturalmente molto simili e si compongono di: segmento esterno: dischi che portano il pigmento visivo soma: con nucleo, mitocondri, reticolo endoplasmico, Golgi etc. segmento interno: terminale sinaptico in contatto con le cellule bipolari.
Struttura di un bastoncello
La parte più caratteristica dei fotorecettori è sicuramente il segmento esterno. In entrambi i casi troviamo una serie di dischi impaccati che derivano dalla membrana esterna del fotorecettore e che, nel caso dei bastoncelli, si distaccano completamente restando isolati a formare una pila di dischi, mentre nel caso dei coni mantengono connessione con la membrana esterna. Questi dischi sono soggetti ad un rapido turnover. Quelli più vecchi sono eliminati all’apice del fotorecettore, mentre quelli più nuovi derivano dalla parte a contatto con il soma.
La ragione per cui questi dischi sono così numerosi è che su di essi è localizzato il pigmento visivo, cioè la sostanza fotosensibile. Questi pigmenti sono così fittamente impaccati sui dischi che costituiscono circa il 40% dell’intera massa del segmento esterno. Questi pigmenti sono proteine coniugate e sono incorporate nei dischi come proteine transmembrana (vedremo che appartengono anch’essi alla categoria dei 7 -helixspanning receptors). Vediamo quali sono le differenze fra coni e bastoncelli.
Rod Cells Cone Cells Very sensitive, necessary for night vision. Low sensitivity, specialized for day vision Saturate in daylight Saturate only in most intense light Discs include much more photopigment in their outer segment Fewer discs and less pigment Can detect a single photon of light of suitable wavelength Detects only multiples of photons Sensitive to scattered light Sensitive to narrow axes of light path
n I bastoncelli sono più sensibili dei coni perché per ogni fotone hanno una variazione di potenziale di riposo 6 volte maggiore dei coni. Quindi la sensibilità della retina alla luce è maggiore dove ci sono più bastoncelli e minore dove ci sono soltanto coni
I coni sono molto concentrati nel punto di max acuità visiva, cioè nella fovea e poi diminuiscono mano a mano che ci si allontana. I bastoncelli sono assenti nella fovea e sono invece distribuiti nella retina laterale
n Acuità visiva: si intende la minima distanza fra due punti che vengano chiaramente percepiti come punti separati ad una distanza standard dell’osservatore.
Rod Retinal Column Cone Retinal Column Poor point resolution Not present in fovea High resolution Concentrated in fovea and evermore defuse periphereally One pigment type. . . no color vision Three pigment types. . . all color vision.
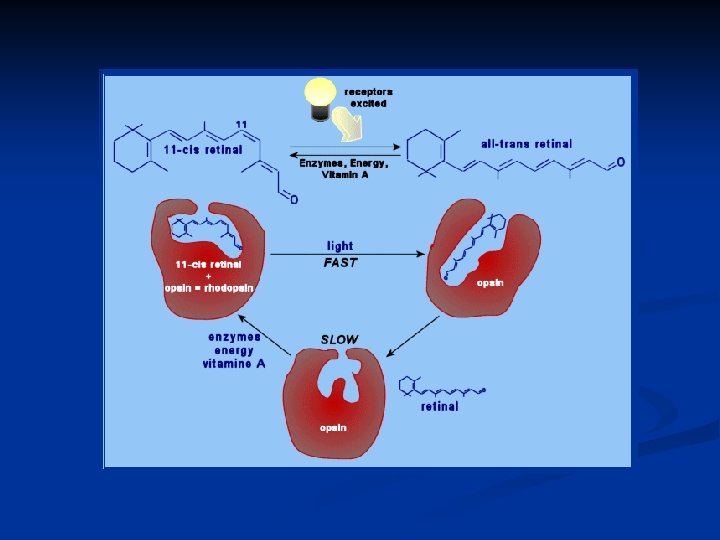
Il pigmento visivo dei bastoncelli prende il nome di rodopsina. Nei coni esistono tre diversi tipi di pigmenti visivi che hanno caratteristiche simili alla rodopsina, ma una diversa sensibilità per le radiazioni dello spettro. La rodopsina è formata da: una parte proteica detta opsina un gruppo cromoforo detto retinale (aldeide della vitamina A) nella conformazione 11 -cis al buio e all-trans sotto stimolazione luminosa.
Quando la rodopsina interagisce con un fotone, la prima reazione che avviene, e fra l’altro l’UNICA luce-dipendente, è l’isomerizzazione del retinale dalla forma 11 -cis alla forma all-trans.
Questo ha come conseguenza immediata il fatto che l’opsina perde affinità per il retinale, perché si viene a creare una sorta di ingombro sterico. La rodopsina passa RAPIDISSIMAMENTE attraverso una serie di intermedi altamente instabili, che sono: prelumirodopsina o batorodopsina lumirodopsina metarodopsina II: è la forma attiva del recettore che determina variazioni elettriche nel fotorecettore
La metarodopsina II subisce un idrolisi del legame fra opsina e all-trans retinale e la molecola di rodopsina si divide. Il pigmento verrà poi ricostituito a livello dell’epitelio pigmentato grazie all’azione di una retinal-isomerasi che isomerizza l’all-transretinale in 11 -cis-retinale che si legherà nuovamente all’opsina riformando il pigmento visivo.
Il ciclo del pigmento visivo
Vediamo adesso come la formazione della metarodopsina II determina una risposta elettrica del fotorecettore.
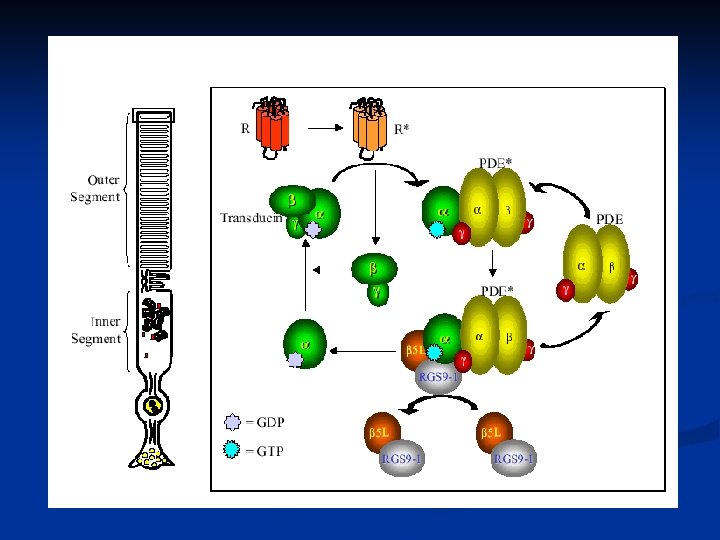
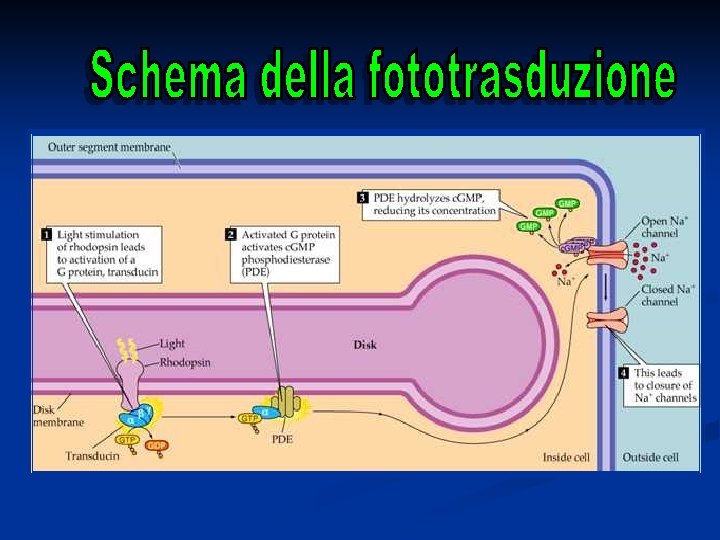
Genesi del potenziale di recettore La rodopsina è un recettore che appartiene alla famiglia dei 7 -helix-spanning receptors (come i recettori -adrenergici e altri) accoppiati a Gproteine. Quando il recettore è attivato, e cioè nel nostro caso si ha la formazione di metarodopsina II, la proteina G viene attivata, iniziando il suo ciclo. La proteina G del ciclo visivo prende il nome di TRANSDUCINA.
È a questo stadio che viene attivata la proteina G e quindi il ciclo visivo
Nel ciclo visivo l’attivazione della proteina G ha lo scopo di attivare un enzima citosolico che si trova nel citoplasma del segmento esterno del bastoncello, che è la fosfodiesterasi (PDE). Questo enzima, una volta attivato catalizza la seguente reazione di deciclizzazione: c. GMP
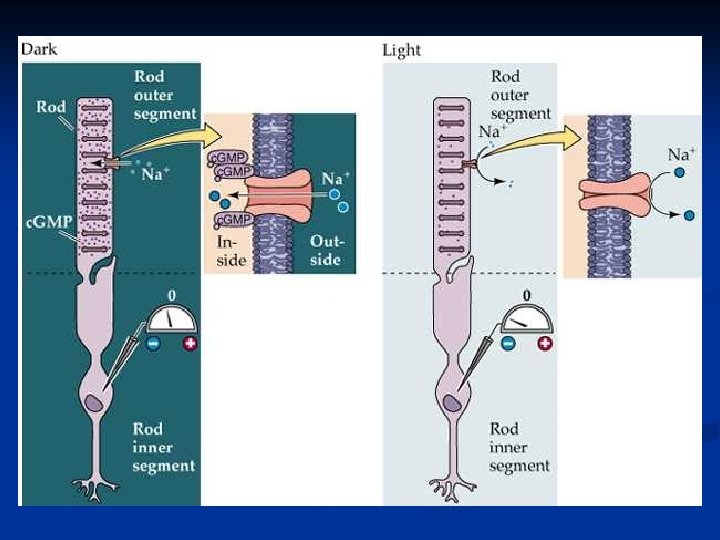
Il risultato dell’attivazione della rodopsina è quindi la diminuzione di c. GMP nel citoplasma (scende sotto a 2µM). Perché il c. GMP deve diminuire? In un fotorecettore al buio la concentrazione del c. GMP è circa 2 M e ha la funzione di MANTENERE APERTI CANALI CATIONICI PRESENTI SULLA MEMBRANA ESTERNA DEL FOTORECETTORE. Questi canali sono permeabili al Na+ e al Ca 2+.
Nel segmento interno ci sono i canali del K+ che, di solito, determinano il potenziale di riposo di membrana. I canali cationici mantenuti aperti dal c. GMP nel segmento esterno, tendono a spostare il potenziale di riposo verso valori meno negativi perché si stabilisce un equilibrio fra canali del K+ e canali cationici. Tanto che IL POTENZIALE DI RIPOSO DI UN FOTORECETTORE AL BUIO VALE CIRCA -40 m. V.
Quando il fotorecettore è stimolato dalla luce e si innesca il ciclo visivo, la PDE determina una diminuzione di c. GMP e quindi i canali cationici si chiudono. I canali K+ diventano quindi i soli determinanti del potenziale di membrana, che diviene più negativo. I FOTORECETTORI STIMOLATI RISPONDONO QUINDI CON UNA VARIAZIONE DEL POTENZIALE DI MEMBRANA IN SENSO IPERPOLARIZZATO RISPETTO AL RIPOSO.
Rods and cones have leaky cell membranes with a continuing inflow of sodium ion then with a standing generator potential. As a result they are continuously releasing their neurotransmitter (glutamate) onto the post-synaptic membrane of the bipolars. If the light receptor is illuminated the bipolar is freed from its ongoing inhibition and depolarizes (generator), causing release of its transmitter (glutamate) onto its ganglion cell.
Nella cascata di eventi appena vista, va notato il fatto che il sistema visivo è in grado di amplificare moltissimo il segnale, fino ad arrivare alla detection anche di un singolo fotone. Questo è possibile grazie al fatto che ciascun evento coinvolto nella cascata della rodopsina determina una notevole amplificazione del sistema. Vediamo cosa significa.
Fotone Un e- dell’ 11 -cis-retinale Transducina PDE. . . . . Transducina PDE Idrolisi di c-GMP . . Idrolisi di c-GMP Canale del Na+ . . Canale del Na+
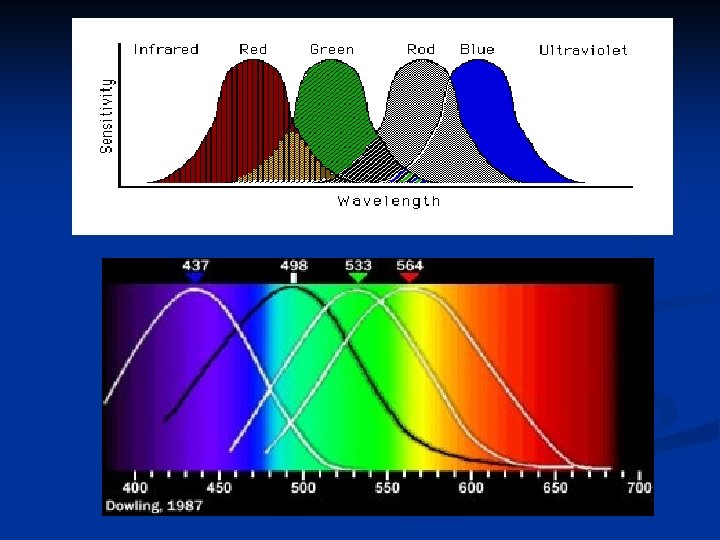
Questo meccanismo a cascata rende i bastoncelli particolarmente sensibili al buio. I coni sono circa 300 volte meno sensibili dei bastoncelli, ma i loro pigmenti funzionano in maniera simile. La differenza dei pigmenti visivi è nella parte proteica, cioè nelle opsine (fotopsine), mentre la parte del retinale è uguale. Esistono quindi tre diverse popolazioni di con tre diversi fotopigmenti che risultano sensibili al blu, al verde e al rosso con picchi di assorbanza a 445, 535 e 570 nm rispettivamente.
L’eccitamento di un certo tipo di recettore permette di sperimentare la sensazione di un certo colore. Gli altri colori dipendono dalla simultanea eccitazione di più di uno di questi recettori. Questa teoria è chiamata teoria tricromatica o teoria di Young-Helmholtz.
Le teorie per spiegare la percezione dei diversi colori sono molte, ma la più accreditata si basa sul fatto che miscelando opportunamente in differenti combinazioni le luci monocromatiche rossa, verde e blu, si ottengono tutti i colori. Per esempio una luce arancio determina un grado di stimolazione dei: coni per il rosso al 99% coni per il verde al 42% coni per il blu al 0% 99 : 42 : 0
Similmente si ha per il giallo che determina una stimolazione delle popolazioni di coni di: coni per il rosso al 83% coni per il verde al 83% coni per il blu al 0% 83 : 0 La percezione del bianco deriva da una stimolazione pressappoco simile per le tre popolazioni di coni.
Quando l’occhio è sprovvisto di uno dei tre tipi di coni per la visione cromatica, viene a mancare la capacità di distinguere certi colori da altri. Per esempio la cecità cromatica per il rosso-verde è un’anomalia genetica che si trasmette attraverso le femmine e colpisce soprattutto i maschi.
Il daltonismo Fu il chimico inglese John Dalton a dare, nel 1794, una descrizione scientifica del daltonismo, pubblicando l'articolo intitolato "Extraordinary facts relating to the vision of colors" (Fatti straordinari legati alla visione dei colori), dopo essersi reso conto della propria cecità cromatica. Il tipo di daltonismo di cui Dalton era affetto oggi prende il nome di deuteranopia.
Esistono diversi tipi di daltonismo. I più diffusi sono dovuti ad alterazioni ereditarie dei fotorecettori, ma è anche possibile diventare daltonici in seguito ad un danneggiamento della retina, del nervo ottico o di determinate aree della corteccia cerebrale. Tale daltonismo è spesso diverso da quello di origine genetica; ad esempio, può manifestarsi solo in una parte del campo visivo. Alcune forme di daltonismo acquisito sono reversibili. Alcune forme temporanee di daltonismo affliggono raramente chi soffre di emicrania.
Visione dicromatica: protanope: insensibile alla luce rossa deuteranope: insensibile alla luce verde Visione monocromatica: totale cecità cromatica
Both normal and those with all colour vision deficiencies should read the number 12.
Those with normal colour vision should read the number 8. Those with red-green colour vision deficiencies should read the number 3. Total colour blindness should not be able to read any numeral.
Normal vision should read the number 29. Red-green deficiencies should read the number 70. Total colour blindness should not read any numeral
Normal colour vision and those with total colour blindness should not be able to read any number. The majority of those with red-green deficiencies should read the number 5.
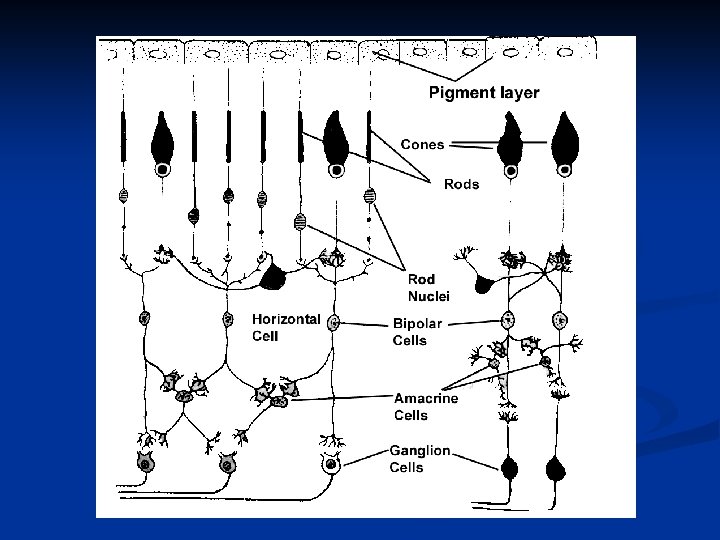
Schema delle connessioni retiniche
Le principali cellule e connessioni nervose che troviamo nella retina sono: Fotorecettori: coni e bastoncelli che sono in contatto con cellule orizzontali o cellule bipolari. Cellule orizzontali: trasmettono segnali orizzontalmente nello strato plessiforme esterno dai bastoncelli e coni ai dendriti delle cellule bipolari
Cellule bipolari: trasmettono segnali dai fotorecettori e cellule orizzontali allo strato plessiforme interno dove sono in sinapsi con cellule amacrine e cellule gangliari. Cellule amacrine: trasmettono segnali orizzontalmente fra i neuriti delle bipolari e i dendriti delle gangliari o di altre cellule amacrine. Cellule gangliari: trasmettono segnali in uscita al nervo ottico.
È importante notare alcune differenze nelle connessioni di coni e bastoncelli. I coni che si trovano nella fovea si connettono 1: 1 con le cellule bipolari, che a loro volta si connettono 1: 1 con le gangliari e da questo dipende l’elevata acuità visiva della fovea. Nasal periphery Al di fuori della fovea sia i coni che i bastoncelli tendono a convergere sulle bipolari.
Nella retina, così come per altri sistemi sensoriali, esistono due sistemi visivi di cui uno legato ai bastoncelli, filogeneticamente più antico e uno legato ai coni, più recente. Le fibre nervose che conducono segnali legati ai coni sono vie più grosse e a maggior velocità di conduzione. Le connessioni retiniche sono riportate schematicamente di seguito.
CONI inibitoria ORIZZONTALI BIPOLARI AMACRINE GANGLIARI Via diretta Via indiretta inibitoria Via indiretta
BASTONCELLI ORIZZONTALI BIPOLARI AMACRINE GANGLIARI N. B. Nel sistema dei bastoncelli la via diretta passa quasi sempre attraverso le cellule amacrine
La trasmissione della maggior parte dei segnali avviene nella retina per conduzione elettrotonica, non per potenziali d’azione. Gli unici neuroni retinici che trasmettono segnali visivi sempre mediante spikes sono le cellule gangliari. Caratteristiche sono le cellule orizzontali, che mediano la risposta fra fotorecettori e bipolari. I segnali in uscita delle cellule orizzontali sono sempre segnali inibitori.
La funzione delle cellule orizzontali è quella di costituire un sistema di inibizione laterale il cui principio fondamentale è quello di esaltare la risposta di un recettore inibendone la periferia. Questo concetto è stato spiegato da Hartline e colleghi nel 1956. In neurobiologia l’inibizione laterale è la capacità che ha un recettore e/o un neurone di ridurre l’attività delle cellule vicine. Questo meccanismo consente intanto di identificare meglio la provenienza dello stimolo e poi di esaltare la risposta di un recettore inibendo la risposta dei recettori vicini. Nel sistema visivo in particolare vuol dire migliorare il contrasto di un’immagine.
Il meccanismo di inibizione laterale è dovuto all’azione delle cellule orizzontali e delle cellule bipolari che determinano quello che viene definito effetto centro-periferia. Il nostro intero sistema visivo esiste per vedere margini e contorni. Noi vediamo il mondo che ci circonda come un insieme di linee. Valutiamo colori e luminosità in base a confronti e NON SECONDO UNA SCALA ASSOLUTA.
Il sistema di inibizione laterale nella retina è il primo passo nel definire e modellare i contorni fra zone chiare e zone scure. La luce diffusa è ignorata dalle cellule gangliari che sono invece attivate da uno spot luminoso ben netto. A livello di corteccia tutti questi punti sono combinati in linee che verranno a loro volta combinati in curve e così via fino alla percezione finale dell’oggetto.
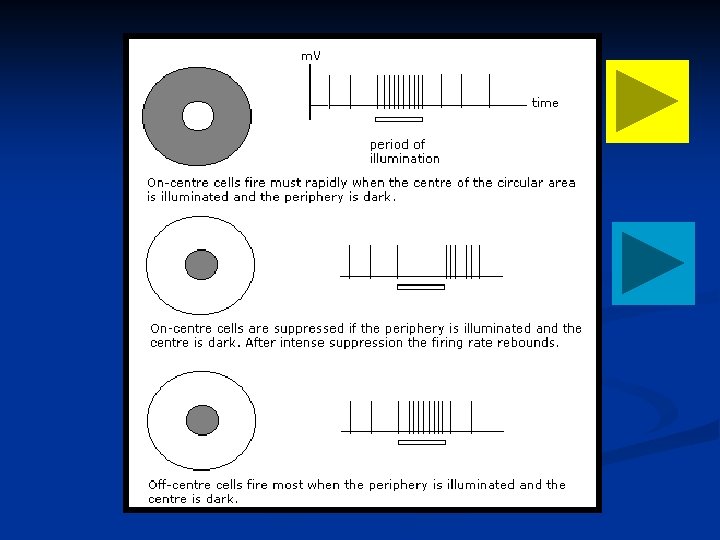
1. La luce iperpolarizza il bastoncello. Diciamo che determina uno stato ON che eccita la bipolare subito sotto. Questa bipolare poi eccita la B gangliare. Lo stesso accade ai fotorecettori vicini. G
2. Il trucco sta nel fatto che le cellule vicine attivano le cellule orizzontali, le quali vanno ad inibire la cellula bipolare centrale sottostante. In questo modo cosa fa la luce diffusa? La cellula bipolare è stimolata, ma anche inibita lateralmente. Il risultato è che la gangliare non viene eccitata e mantiene il suo stato tonico di scarica
3. Un piccolo spot centrale di luce tuttavia eccita il fotorecettore e la bipolare, ma non le cellule vicine. Non c’é inibizione laterale e quindi la bipolare riesce ad eccitare a sua volta la gangliare che aumenta la sua frequenza di scarica.
4. Un anello di luce eccita solo i fotorecettori vicini. La bipolare ora è fortemente inibita. In risposta a questo deciso silenzio della bipolare, la gangliare riduce la sua scarica e non si riattiverà fino a quando la luce non viene a mancare sui fotorecettori vicini
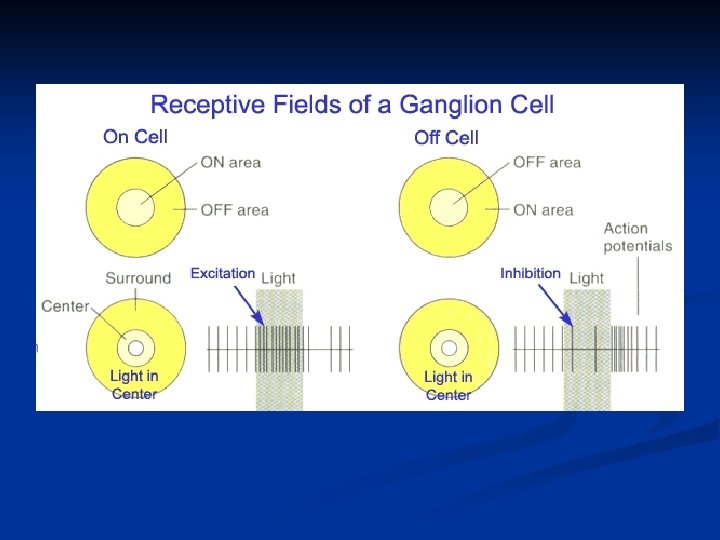
Questa viene definita cellula bipolare a centro ON. L’opposto di questo scenario può essere creato invertendo tutti i segnali con il risultato di avere una cellula a centro OFF. Questa peculiarità di centro ON / OFF è anche una caratteristica dei neuroni del nucleo genicolato laterale talamico. Le cose divengono ancora più complesse a livello corticale.
Cellule bipolari ON e OFF Esistono due diversi tipi di cellule bipolari: le bipolari a centro ON e le bipolari a centro OFF. Le prime depolarizzano quando il fotorecettore con cui sono in contatto è eccitato, mentre le altre iperpolarizzano. Siccome le bipolari ON e le bipolari OFF sono adiacenti, questo costituisce un sistema molto raffinato per la percezione dei contrasti anche quando il fronte di contrasto cade fra due recettori adiacenti.
Centro ON Periferia OFF Inibizione
Una cellula bipolare ON è in sinapsi con una gangliare ON e lo stesso per le bipolari OFF ON center OFF center
Qual’è il vantaggio di un antagonismo centro-periferia? Questo contrasto aiuta il sistema visivo a percepire i bordi degli oggetti. Questi si distinguono dalla periferia per il brusco cambiamento di riflessione della luce al loro contorno. In questo modo la percezione dei contrasti aiuta a definire il contorno delle cose distinguendole dallo sfondo. Aree delle immagini che non cambiano contrasto o lo cambiano poco, probabilmente appartengono allo stesso oggetto. Il sistema visivo è sostanzialmente interessato soltanto a individuare aree che cambiano luminosità, non ai valori assoluti.
Mach band effect
Abbiamo visto che le vie più semplici prevedono un processamento dell’informazione visiva fra coni, bipolari e gangliari e fra bastoncelli, bipolari, amacrine e gangliari. In realtà le connessioni sono molto più complesse perché prevedono anche l’azione integrata con cellule orizzontali e amacrine. Di seguito è riportato un grafico indicativo delle possibili relazioni che intercorrono fra i neuroni retinici.
La retina di ciascun occhio contiene circa 120 milioni di bastoncelli e 6 milioni di coni, mentre il numero delle gangliari è di circa 1. 5 milioni. Su ogni fibra del nervo ottico si ha quindi convergenza di bastoncelli e coni. Inoltre nella fovea i bastoncelli sono assenti e ci sono circa 35. 000 coni. Questo spiega il notevole aumento di acuità visiva che si ha procedendo verso la fovea.
A livello foveale addirittura si ha un rapporto 1: 1 fra coni e gangliari e la massima acuità visiva. Bisogna ricordare che le cellule gangliari trasmettono i segnali attraverso il nervo ottico mediante potenziali d’azione. Anche le cellule gangliari sono soggette a un meccanismo a centro ON o OFF e questo è particolarmente importante nella percezione dei contrasti.
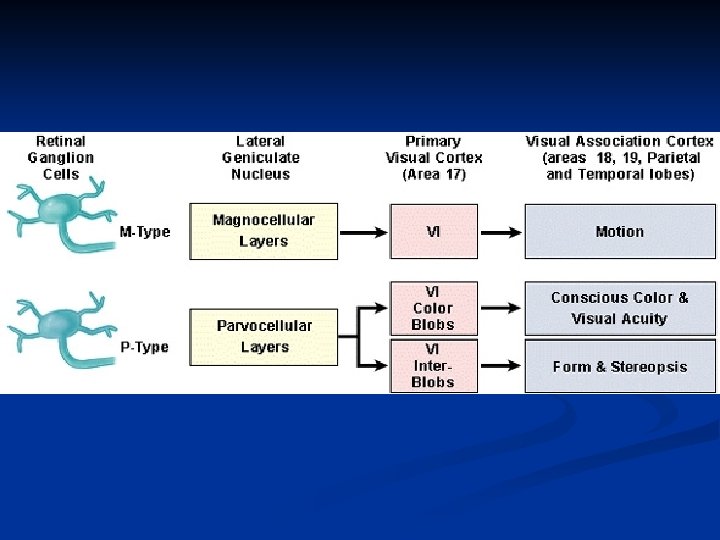
Esistono almeno 18 tipi diversi di cellule gangliari nella retina umana. Fra queste due classi sono tipiche dei primati e sono implicate nell’elevata acuità visiva e nella percezione dei colori. Si tratta delle cellule gangliari P e M che proiettano rispettivamente ai neuroni parvocellulari e magnocellulari del nucleo genicolato laterale del talamo.
Gli impulsi lasciano la retina attraverso il nervo ottico (II). Ogni nervo ottico è formato da circa 1. 5 milione di assoni che, ovviamente occupano spazio, per cui la papilla rappresenta un punto cieco della retina perché non ha spazio per i fotorecettori. Poiché le papille dei due occhi non coincidono nel campo visivo, l’uno complementa l’altro.
The Blind Spot Close your left eye and hold the following page about 8 -9 inches from your right eye. Look at the cross and slowly move the page forward and backward until the black circle disappears. This will happen when light rays from the black circle fall on the place where the optic nerve enters the retina commonly referred to as your "blind spot".
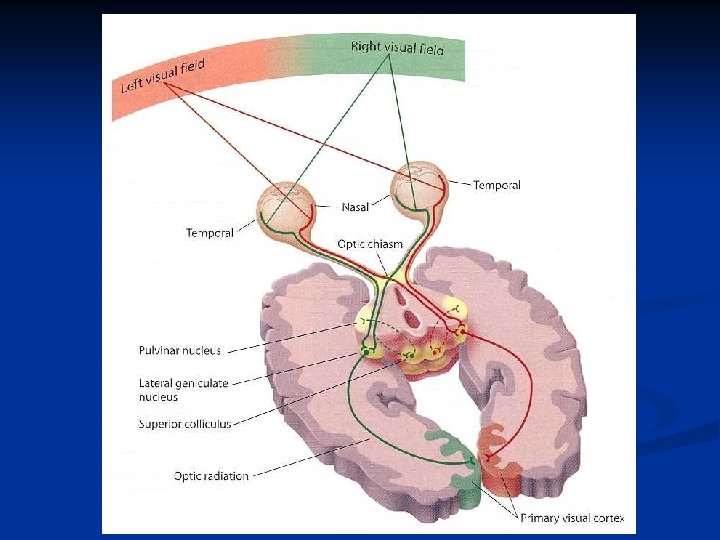
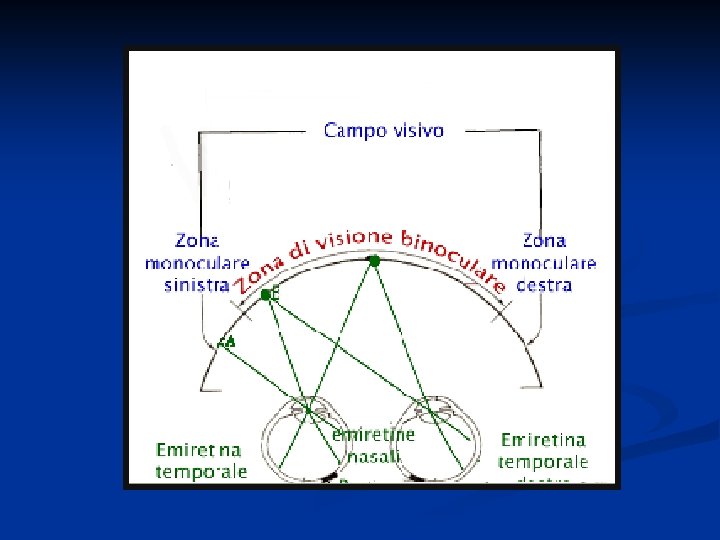
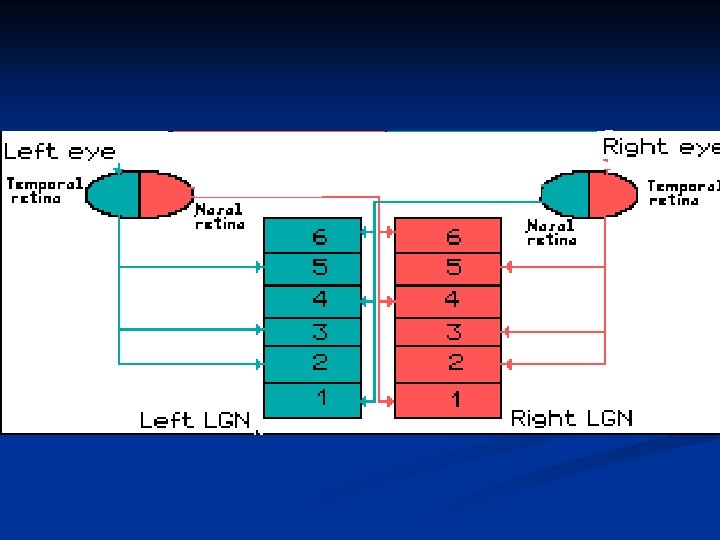
A livello del chiasma ottico tutte le fibre provenienti dalla retina nasale di ciascun occhio si incrociano e passano nel lato opposto dove si uniscono con le fibre temporali della retina di quel lato. Si formano così i tratti ottici.
Il campo visivo è la proiezione ottica della rappresentazione visiva del mondo esterno sulla retina. L'uomo presenta una visione binoculare molto ampia, cosa che gli consente di aver consapevolezza della distanza relativa degli oggetti
Le fibre di ciascun tratto si portano al nucleo genicolato laterale del talamo e da qui il tratto genicolo-calcarino si porta alla corteccia visiva I in zona occipitale che corrisponde all’area 17 di Brodmann. Da questa si passa alle aree di associazione 18 e 19.
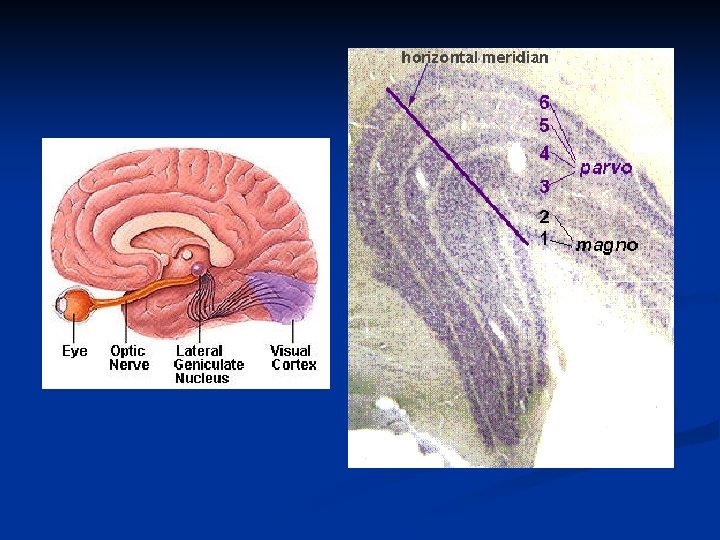
Nucleo genicolato laterale Il n. g. l. è formato da 6 strati di neuroni organizzati in maniera molto precisa. In primo luogo, le cellule dello stesso strato nella parte di dx e nella parte di sx hanno campi recettivi nella stessa area della retina. Inoltre le fibre provenienti dai due occhi sono segregati in strati diversi. Le fibre ipsilaterali arrivano negli strati 2, 3, 5, mentre le fibre controlaterali agli strati 1, 4, 6.
n Tutte le cellule nel n. g. l. hanno campi recettivi concentrici come le cellule gangliari. In particolare gli strati 1 e 2 sono formati da neuroni magnocellulari che mediano la risposta acromatica, mentre gli strati 3, 4, 5 e 6 sono formati da neuroni parvocellulari che mediano la visione cromatica
Occhio Nervo ottico Chiasma ottico Tratto ottico Nucleo genicolato laterale Tratto genicolocalcarino Corteccia I
Fibre visive si dirigono anche ad aree encefaliche più antiche. Dai tratti ottici: § al nucleo soprachiasmatico dell’ipotalamo § ai nuclei pretettali per il riflesso pupillare e la fissazione degli occhi § al collicolo superiore del mesencefalo per i movimenti simultanei due occhi § al nucleo genicolato ventrale per partecipare al controllo delle funzioni comportamentali legate alla visione.
Anche per la corteccia visiva si conosce un area I e una corteccia visiva II. La corteccia visiva I si estende al polo occipitale e qui terminano i segnali visivi provenienti dagli occhi. Punti specifici della retina sono connessi con punti specifici della corteccia. In particolare la rappresentazione della fovea è estesissima rispetto a quella di zone più periferiche della retina. La corteccia I (V-1) corrisponde all’area 17 di Brodmann e comprende la corteccia più spessa di tutto l’encefalo. Ha la funzione di codificare le linee e i confini degli oggetti del campo visivo
17
Le aree visive di associazione sono localizzate anteriormente, superiormente e inferiormente a V 1. Si tratta di aree di associazione indicate da V-2 a V-3, V-4 etc. in cui sono progressivamente selezionati ed analizzati i vari aspetti dell’immagine visiva. L’informazione visiva passa da V-1 alla corteccia secondaria seguendo due canali principali.
Una via serve per l’analisi della posizione tridimensionale, della forma grossolana e del movimento di oggetti (via dorsale, via M alla corteccia parietale). Una seconda via permette l’analisi dettagli visivi e del colore (via antero-laterale, via P alla corteccia infero-temporale).