IL DNA E UNA MOLECOLA DINAMICA STABILITA vs

IL DNA E’ UNA MOLECOLA DINAMICA STABILITA' vs INSTABILITA' GENOMA

DNA REPAIR

Errori da replicazione del DNA • il DNA è replicato in 2 -3 ore • ~100, 000 base pairs sono replicate in 1 secondo • è fatale che accadano errori • se non fosse per il sistema di correzione errori, le mutazioni prodotte sarebbero assai più numerose, probabilmente di un livello insopportabile (!)

Sono decine di migliaia ogni giorno le mutazioni che colpiscono una cellula. Ma solo lo 0, 02% di queste si accumulano nel DNA in modo permanente. Tutte le altre sono eliminate mediante riparazione del DNA. Quanto sia importante questo sistema è dimostrato anche dal fatto che una apprezzabile quota percentuale del genoma codificante è impegnata da geni implicati nel funzionamento del sistema stesso, nell' aumento del tasso di mutazione quando un gene del sistema viene inattivato, dalla diffusione del sistema e dei geni in pressochè ogni organismo vivente


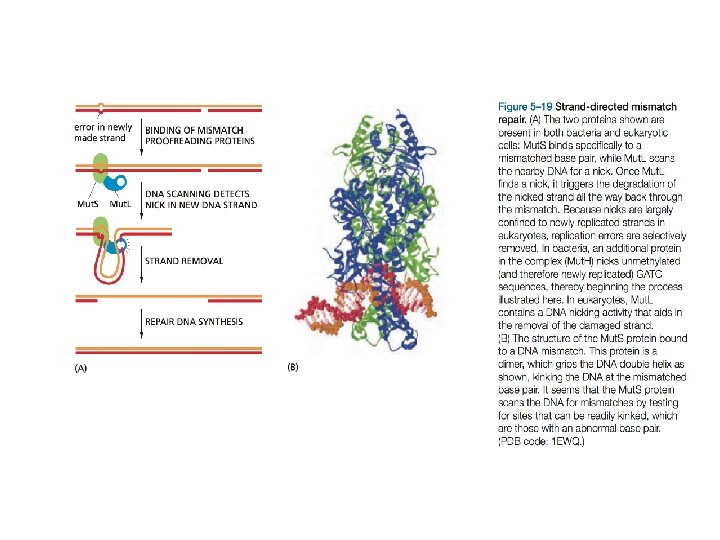
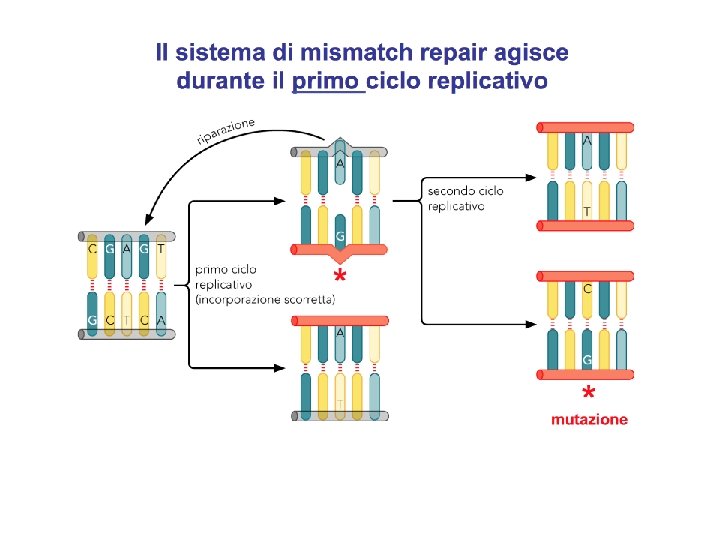
Lo "strand-Directed Mismatch Repair System" rimuove gli errori di replicazione che sfuggono alla "macchina" replicativa

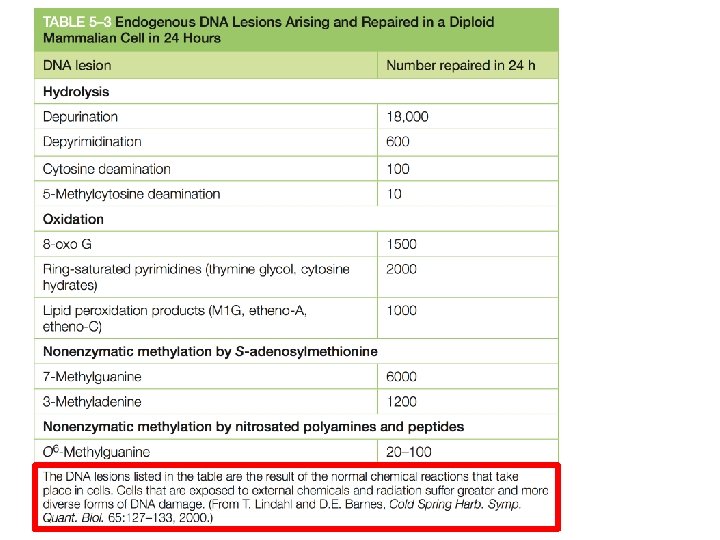
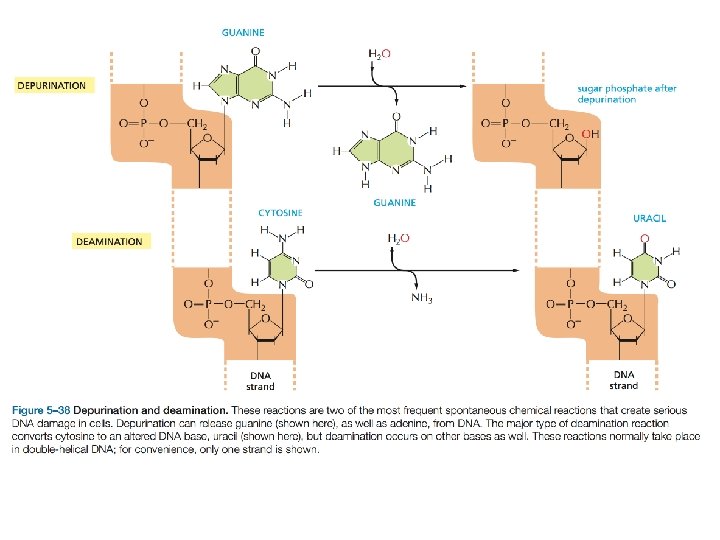
Le più comuni conseguenze sono la delezione di una o più basi oppure la sostituzione di una coppia di basi 18, 000 basi puriniche vengono perse ogni giorno per idrolisi del legame Nglicosidico in ogni cellula. 100 citosine al giorno per cellula subiscono la deaminazione a uracile
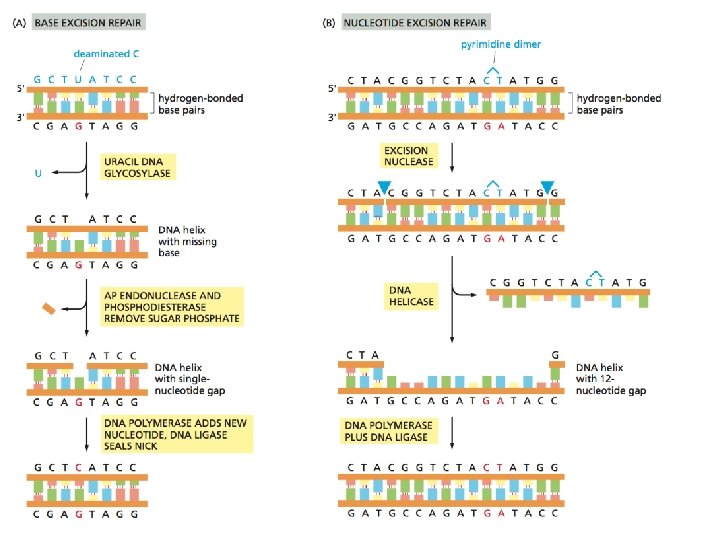

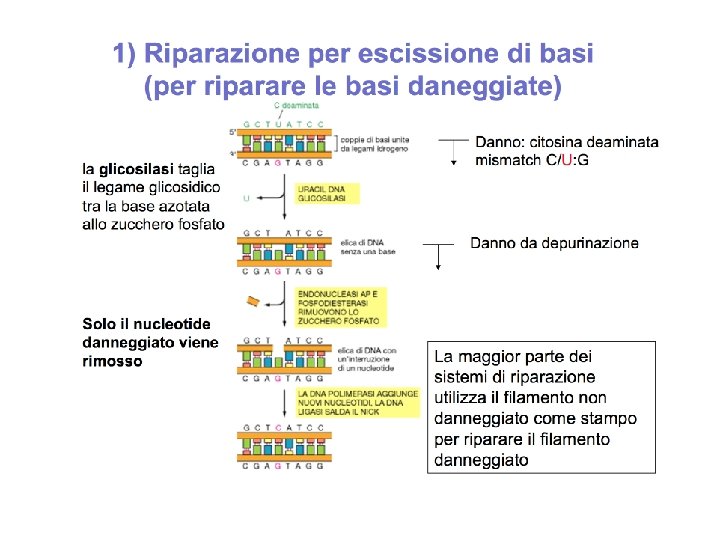
Nel caso di BER una batteria di DNA glicosilasi, ciascuna delle quali riconosce uno specifico tipo di base alterata, catalizza la rimozione idrolitica della base. Vi sono almeno 6 tipi di questi enzimi che rimuovono citosine e adenine deaminate, basi alchilate oppure ossidate, basi con anelli aperti, basi in cui il doppio legame tra due atomi di carbonio è convertito a singolo legame. Come vengono riconosciute queste basi alterate ?

Sembra che per il riconoscimento della base alterata la DNA glicosilasi si muove su e giu lungo la catena del DNA rivoltando le basi verso l' esterno (“flipping-out”). Se la base viene riconosciuta come alterata, allora viene rimossa.
 endonucleasi, che")
Il vuoto lasciato dalla base mancante viene riconosciuto da una AP (APurinic/APyrimidinic) endonucleasi, che taglia la sequenza a livello del legame fosfodiesterico.

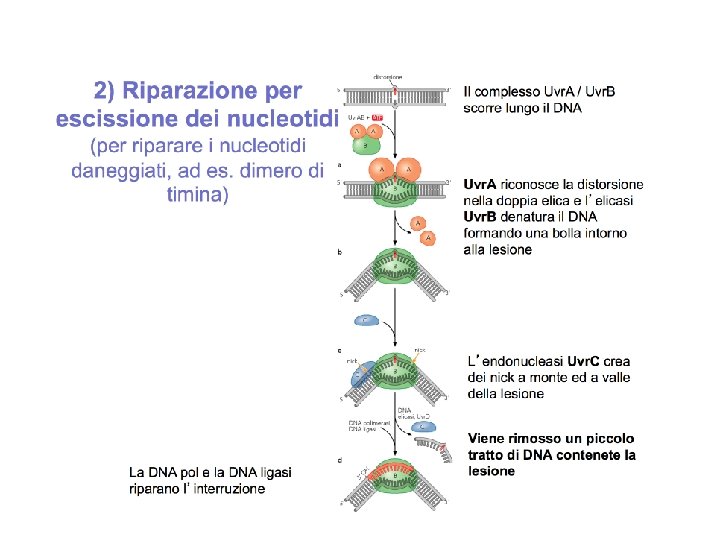
La NER ovvero nucleotide excision repair. E' il meccanismo che ripara il DNA da quasi qualunque tipo di lesione che coinvolge più di un singolo nucleotide. Ad es. reazioni covalenti con composti come benzopyrene, presente nel fumo di tabacco, o i fumi liberati da motori diesel, e inoltre i dimeri di pirimidine (T-T, T-C, and C-C).

Un grosso complesso multienzimatico esamina il DNA per la presenza di distorsioni a livello della doppia elica. In caso positivo la porzione di catena alterata viene tagliata via a livello del legame fosfodiesterico agendo a monte e a valle della lesione. Una DNA elicasi rimuove il frammento tagliato e la riparazione viene completata al solito da una DNA polimerasi e la DNA ligasi

Un meccanismo alternativo è costituito dalla cosidetta "reversione chimica diretta" del danno. Questo meccanismo è impiegato per la rimozione rapida di talune lesioni altamente mutageniche o citotossiche, ad es. la formazione mediante alchilazione di O 6 -methylguanine cui viene rimosso il gruppo metilico mediante trasferimento diretto a un residuo di cisteina presente sulla medesima proteina che effettua la riparazione, e che viene poi eliminata

Coupling Nucleotide Excision Repair to Transcription Ensures That the Cell’s Most Important DNA Is Efficiently Repaired

La RNA polimerasi si blocca a livello della lesione e richiama il sistema di riparazione per excisione. I pazienti affetti da Cockayne syndrome presentano una associazione difettosa tra NER e trascrizione. Essi presentano ritardo dello sviluppo, alterazioni scheletriche, ritardo progressivo del sistema neuronale, e sensibilità alla luce. Queste alterazioni sono probabilmente conseguenti al blocco della RNA polimerasi nel tentativo di trascrivere la sequenza codificante di importanti geni dello sviluppo

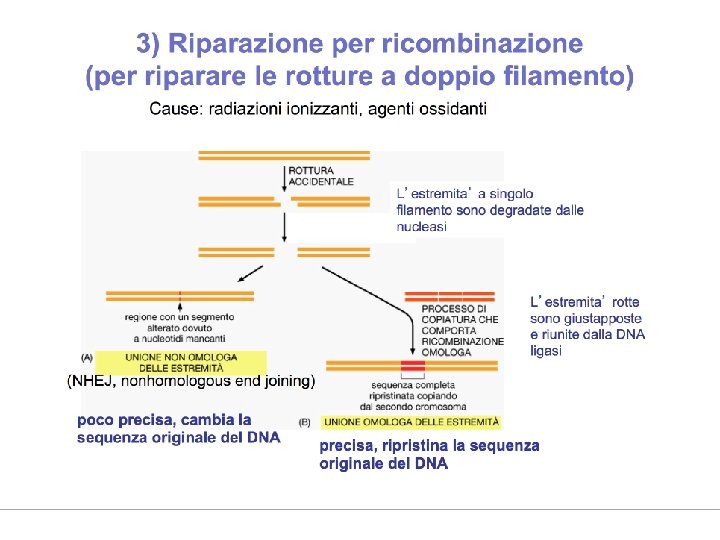
Double-Strand Breaks Are Efficiently Repaired

I meccanismi di riparazione sono diversi per questo tipo di lesione. Legame delle estremità non omologhe con perdita di nucleotidi a livello delle estremità. Meccanismo veloce ma "sporco", soluzione molto usata dalle cellule somatiche di mammifero. Dà luogo a mutazione, anche se la probabilità che la mutazione sia patogena è molto bassa. Soluzione accettabile al problema di ricucire cromosomi tra loro
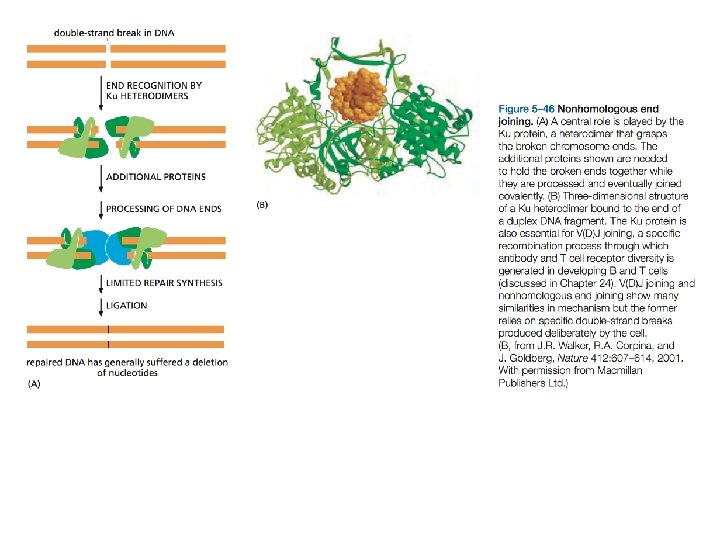

Alla età di 70 anni è possibile avere accumulato i segni di oltre 2000 di tali maldestri tentativi di ricongiunzione di estremità rotte di cromosomi. Ma il rischio mutagenico è in realtà un altro: il rischio che le estremità ricongiunte appartengano a due cromosomi differenti e che si generino quindi cromosomi dicentrici e acentrici che missegregano. Quanto questo rischio sia grave è rappresentato indirettamente dalla evidenza che le estremità naturali dei cromosomi (telomeri) sono rese non "appiccicose", per impedire che siano riconosciute come estremità rotte da riparare.

II meccanismo Si avvale della ricombinazione omologa e dei cromatidi fratelli come stampo. Viene impiegato soltanto in corso di replicazione del DNA quando diventano disponibili i cromatidi

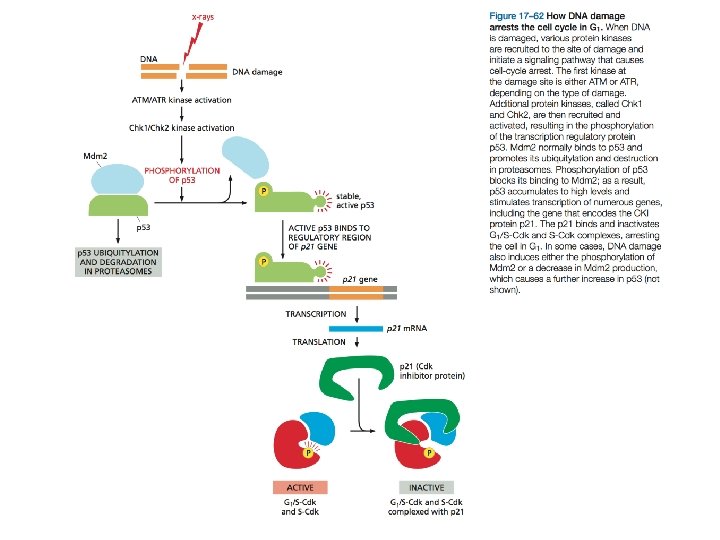
DNA Damage Delays Progression of the Cell Cycle

Va detto in conclusione che la risposta al danno nel DNA è mediata da una serie di messaggeri che segnalano la presenza del danno e up-regolano la sintesi degli enzimi riparatori. Uno di questi segnalatori è ATM, la cui assenza per mutazione inattivante è responsabile della atassia telangectasica. I sintomi comprendono neurodegenerazione, instabilità genomica e predisposizione ai tumori.

Diverse malattie sono associate a difetti nel riparo del DNA
 defects in")
XP Cockaine syndrome Trichothiodystrophy defects in nucleotide excision repair HNPCC (Lynch syndrome) defects in DNA mismatch repair Ataxia-Telangiectasia defect in repair of radiation induced DNA damage, ATM kinase defect in cell cycle control Fanconi anemia impaired in the removal of interstrand crosslinks (Myto. C), defects in helicase, rischio di leucemia Bloom syndrome defects in helicase Nijmegan breakage syndrome defects in helicase Rothmund-Thomson syndrome defects in helicase Werner syndrome defects in helicase Sono tutti processi a trasmissione autosomico-recessiva

Raggi UV causano Dimeri di Timina Disponiamo di efficienti sistemi di riparo per correggere i dimeri: fotoliasi + luce bianca revertono il dimero alla forma normale (fotoriattivazione) Xeroderma pigmentoso fase tardiva

I due meccanismi di riparo di dimeri di timina Nel riparo mediante fotoriattivazione, l'enzima PRE con il concorso della luce visibile spezza il dimero e rinormalizza l'appaiamento delle basi. Nel riparo da excisione, il sistema genico UVR excide il dimero e rimpiazza i nucleotidi mancanti con l'aiuto della DNA Polimerasi I
 (OMIM 278700 etc. ) Pelle estremamente secca (xeroderma) e lesioni puntiformi")
Xeroderma Pigmentosum (XP) (OMIM 278700 etc. ) Pelle estremamente secca (xeroderma) e lesioni puntiformi maligne Morte prematura di cellule nervose (aging ? ) Il soggetto a sx è stato esposto al sole e ha sviluppato le lesioni Il soggetto a dx ha evitato la esposizione XP insorge per difetto in almeno 8 geni differenti sette dei quali sono necessari nella attività di riparo del DNA per excisione XPA XPB ……XPG

Cockayne syndrome e tricothiodystrophy sono associate a difetti nella attività excinucleasica associata a trascrizione Sono affetti da ritardo mentale mentre i pazienti con tricothiodystrophy hanno bassa statura, capelli fragili e cute di tipo squamoso

HNPCC è associata a difetti del sistema di riparo dei difetti da mismatch. Può essere provocata da difetti in almeno 7 geni differenti. Alcuni di questi geni hanno omologhi in E. coli e S. cerevisiae sottolineando quanto comuni sono le vie di riparo del DNA presenti in organismi tanto differenti. Per quanto riguarda AT, Fanconi, Bloom, Werner, Rothmund. Thomson, Nijmegan: presentano alto rischio per tumori in particolare leucemie AT presenta una elevata sensibilità alle radiazionizzanti suggerendo un difetto nel meccanismo di riparo da i danni provocati da tale tipo di radiazioni. E’ provocata da difetti in una kinase coinvolta nel controllo del ciclo cellulare

In AF incapacità di rimuovere i legami crociati tra i filamenti di DNA che si formano in seguito ad esempio il trattamento con Mitomicina C Bloom e Nijmegan presentano elevata frequenza di rotture cromosomiche esitano in aberrazioni cromosomiche e scambi tra cromatidi fratelli


FINE


: HO 2")
Le difese dell' ospite nei riguardi del danno ossidativo superoxide dismutase (SOD): HO 2 - + H H 2 O 2 catalase: H 2 O 2 H 2 O
- Slides: 46