I MERISTEMI DELLAPICE DEL GERMOGLIO E DELLA RADICE




aumentano la circonferenza di fusti e radici")


")








 PIN espresse in maniera asimmetrica: ØLocalizzazione")
 Espressi con diversa localizzazione in tempi")







 Considerato un gene associato alle cellule iniziali, marcatore dell’attività meristematica dell’apice caulinare")
")

 ØI geni CUC sono necessari")



 conservato helix-turn-helix di binding al DNA di 180 bp")

 identificata originariamente nel mais Mutazione gain of function (dominante)")

")
 KNAT 2 KNAT 6")



: STM espresso specificamente nelle cellule che diverranno cellule meristematiche Mutanti stm")





 che")
 (gene homeobox di classe wox) espresso nel centro organizzatore (OC) sotto la")
 E CLAVATA (CLVI, CLV")

 CLAVATA 2 manca del dominio chinasico")













- Slides: 70
I MERISTEMI DELL’APICE DEL GERMOGLIO E DELLA RADICE
Meristemi: popolazioni di cellule piccole, isodiametriche, con caratteristiche embrionali da poche centinaia a migliaia di cellule (SAM arabidopsis 60) I meristemi vegetativi oltre a generare gli organi della pianta si rigenerano; alcune cellule infatti non si differenziano ma mantengono la capacità di dividersi Le cellule indifferenziate si chiamano INIZIALI (simili alle staminali animali) (stem cells)
ØSi formano durante l’embriogenesi ØPiù corretto il termine protomeristemi ØIl protomeristema apicale è riconoscibile dallo stadio a cuore (le cellule interposte tra i cotiledoni si dividono in maniera orientata e si stratificano) ØProbabilmente l’identità di cellule meristematiche apicali è acquisita più precocemente (stadio globulare) ØLe iniziali della cuffia radicale si formano dall’ipofisi nello stadio a cuore,
Meristemi primari: generano il corpo primario della pianta, dopo la germinazione, si formano durante l’embriogenesi Meristemi ascellari: si formano alle ascelle delle foglie; derivano dal meristema apicale Meristemi delle radici laterali: struttura di meristemi primari si formano dalle cellule del periciclo
Meristemi secondari I meristemi secondari (o laterali )aumentano la circonferenza di fusti e radici producendo tessuto vascolare secondario e periderma Ø Cambio vascolare: iniziali fusiformi e iniziali dei raggi; xilema e floema secondario; raggi di tessuto parenchimatico ØCambio subero fellodermico: si sviluppa nelle cellule mature del cortex e del floema secondario; si differenzia in cellule del sughero che costituiscono il periderma
Meristemi fiorali: derivano dai meristemi vegetativi, producono gli organi fiorali, sono a crescita DETERMINATA anziché indeterminata Ø Meristemi delle infiorescenze: derivano dai meristemi vegetativi e producono brattee e meristemi fiorali alle ascelle delle brattee; possono essere determinati o indeterminati
Crescita secondaria nel fusto
STRUTTURA DEL GERMOGLIO gemma apicale: primordi fogliari, meristema apicale (doma)
Meristema apicale del germoglio ØA crescita indeterminata Øgenera il fusto e gli organi laterali ad esso attaccati: foglie e gemme laterali Ø Contiene diversi strati e zone funzionali Fitomero: internodo con foglia e gemma ascellare
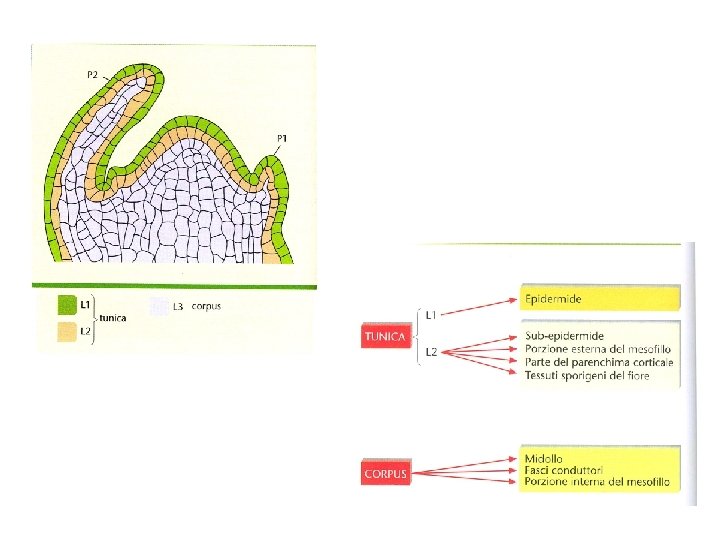
L 3 corpus L 1, L 2 tunica Aspetto stratificato: tre strati L 1; L 2; L 3
L 1 strato più esterno L 1 e L 2 : divisioni anticlinali L 3: piano di divisione meno orientato Tutti e tre gli strati hanno le proprie cellule iniziali e tutti e tre concorrono alla formazione del fusto e degli organi laterali
Il doma ha anche un’organizzazione radiale Ogni zona composta da cellule distinguibili non solo per il piano delle divisioni ma anche per differenze in dimensioni, grado di vacuolizzazione. Hanno pattern diversi di espressione genica che riflettono le differenze di funzione tra le varie zone Zona Centrale: iniziali apicali ; divisioni lente Zona periferica: primordi fiorali; divisioni rapide Costa meristematica: tessuti interni del fusto
Zonazione radiale del doma cellule derivate che si differenziano
FORMAZIONE DEL MERISTEMA APICALE DEL GERMOGLIO
Trasporto di IAA in relazione alla formazione del meristema apicale
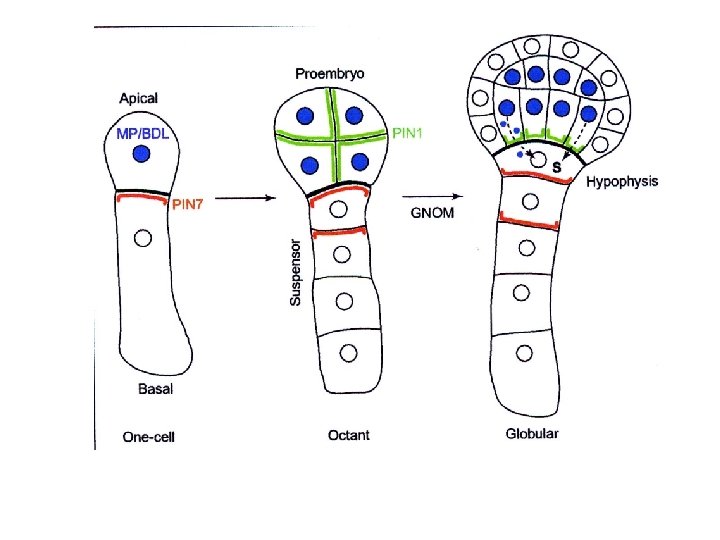
Il meristema apicale del germoglio si forma in una zona di bassa concentrazione di auxina Nell’embrione vengono espressi i carriers PIN 1 (PIN 3) PIN 4 e PIN 7
PIN 1; PIN 4; PIN 7; (PIN 3) PIN espresse in maniera asimmetrica: ØLocalizzazione correla con la direzione del trasporto di IAA ØPIN riciclate continuamente tra reticolo e membrana plasmatica richieste proteine ARF-GEF (GNOM) ØNon noto se PIN di per se carrier o in associazione con ABC transporter (MDR-like)
PIN 1; PIN 4; PIN 7; (PIN 3) Espressi con diversa localizzazione in tempi diversi dello sviluppo dell’embrione La sequenza di espressione regolata temporalmente e spazialmente, è responsabile della variazione nella direzione del flusso di IAA durante l’embriogenesi Nello stadio precoce il flusso di IAA è verso l’apice, lontano dal sospensore; dallo stadio globulare tardivo il flusso è invertito, verso l’ipofisi e la radice in sviluppo. PIN 7, PIN 1 i primi geni PIN espressi nell’embrione: PIN 7 si trova nella membrana apicale della cellula basale nell’embrione allo stadio a due cellule e nella membrana apicale delle cellule del sospensore fino allo stadio a 32 cellule Trasporto di IAA da cellule basali verso l’apice
PIN 1: espresso in maniera non polare nell’embrione fino allo stadio a 32 cellule; Successivamente si localizza nella parte basale delle cellule del procambio; in questo stadio PIN 7 si localizza nella parte basale delle cellule del sospensore e PIN 4 si accumula nella ipofisi e nelle iniziali vascolari Il flusso di IAA è invertito verso l’ipofisi e il sospensore (PIN 3 espresso nel polo radicale nello stadio a cuore)
Nel complesso l’azione combinata di PIN 1; PIN 4; PIN 7 serve a invertire il flusso di IAA nello stadio globulare dall’embrione propriamente detto verso il polo basale
La funzione dei geni PIN è RIDONDANTE ØSingoli mutanti pin hanno fenotipo debole e alla fine sviluppano embrioni quasi normali ØDoppi o tripli mutanti pin hanno alterazioni dello sviluppo più marcate ØMutanti gnom hanno un fenotipo molto marcato Il gene GNOM determina il riciclo e la rilocalizzazione delle proteine PIN e presumibilmente mutazioni gnom influenzano il riciclo di tutti i componenti della famiglia PIN
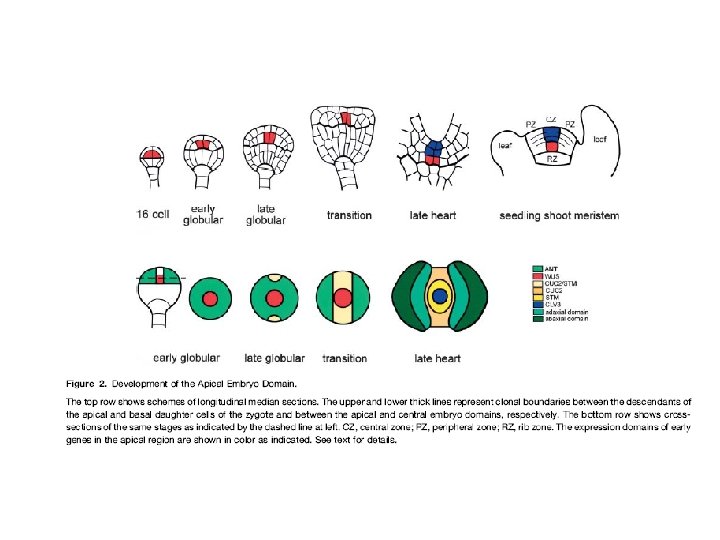
La formazione del meristema apicale dipende dall’espressione di geni modulati da IAA ØNell’embrione precoce i valori di IAA sono elevati e ciò mette in moto il programma di patterning assiale con l’espressione dei geni MONOPTEROS (MP; NPH 4) ØAllo stadio globulare tardivo, la rilocalizzazione delle proteine PIN fa si che il flusso di IAA sia diretto verso le zone fiancheggianti (cotiledoni) la zona centrale dove si formerà il meristema ØLa zona centrale diventa povera di IAA, l’espressione dei geni MP è più debole che nelle zone cotiledonari ØLa ridotta espressione dei geni MP consente l’espressione dei geni CUC (nella zona centrale) necessaria per la determinazione dell’identità del meristema.
ØLa formazione del meristema apicale e della zona intercotiledonaria è correlata alla espressione dei geni CUC 1 CUC 2 e CUC 3 ØI geni CUC vengono espressi nella zona intercotiledonaria dove reprimono la crescita; crescita che ha luogo nelle zone cotiledonari, dove alti livelli di IAA reprimono l’espressione dei geni CUC e determinano il passaggio alla simmetria bilaterale dello stadio a cuore
Formazione del meristema apicale e dei cotiledoni Meristema apicale cotiledoni Espressione ectopica Zona intercotiledonaria ad ab cuc 1 pin 1 pid
MODELLO IAA è trasportato via dalla regioni intercotiledonaria verso le zone cotiledonarie, dove è in grado di determinare la degradazione di un repressore AUX/IAA Ciò consente a geni del tipo MONOPTEROS (ARF) di reprimere la trascrizione dei geni CUC Differenziamento dei cotiledoni
SHOOTMERISTEMLESS (STM) Considerato un gene associato alle cellule iniziali, marcatore dell’attività meristematica dell’apice caulinare Mutanti stm mancano di doma caulinare e di primordi fogliari ma si sviluppano cotiledoni, ipocotile e radice.
Pattern di espressione di CUC e SHOOTMERISTEMLESS (STM)
ØL’espressione di CUC 1 e 2 ha inizio in patches isolati delle cellule apicali ØSi diffonde ad occupare una striscia intorno all’apice dell’embrione dividendolo in una zona centrale e due zone periferiche ØIl pattern di espressione e la funzionalità di CUC 1 e 2 richiede MP e PIN 1 ØLa funzione CUC è necessaria per l’attivazione di STM nella striscia centrale dell’apice ØSTM a sua volta promuove l’espressione di CUC 1 il quale è necessario per la corretta espressione spaziale di CUC 2 ØNello stadio a cuore STM e CUC 2 assumono pattern di espressione complementari STM nel meristema in formazione CUC 2 ai confini tra il meristema apicale e i cotiledoni
LE INTERAZIONI TRA CUC E STM SONO COMPLESSE (indirette) ØI geni CUC sono necessari per l’espressione di STM (STM non è espresso in doppi mutanti cuc 1/cuc 2) ØSTM è richiesto per la corretta espressione spaziale dei geni CUC (l’espressione di CUC 2 è alterata in mutanti stm) CUC appartengono alla famiglia di fattori di trascrizione che contengono domini NAC: dominio di binding al DNA; N-terminale; esclusivo delle piante; twisted b- sheet circondato da diversi elementi a elica; in arabidopsis 105 geni NAC, in riso 75.
In arabidopsis STM è espresso nel doma insieme ad altri geni KNOX come KNAt 1 e KNAT 2 STM nel doma inibisce l’espressione di ASYMMETRIC LEAVES 1 (AS 1) AS 1 si esprime nei primordi fogliari dove reprime l’espressione di KNAt 1 e KNAT 2
Mantenimento dell’identità delle cellule meristematiche Il meristema apicale, a crescita indeterminata dà luogo a strutture determinate come le foglie tuttavia il mantiene le sue dimensioni perché un numerocostante di cellule rimane nello stato differenziato Geni HOMEOBOX della classe KNOX necessari al mantenimento della identità meristematica STM è un gene KNOX (class I knotted homeobox gene) ortologo di KNOTTED 1 in mais (KN 1)
GENI OMEOTICI Scoperti in Drosophila, estremamente conservati negli Eucarioti In Drosophila determinano l’identità anatomica dei segmenti Mutazione omeotica In Drosophila ANTENNAPEDIA Omeosi: trasformazione di una parte del corpo in un’altra
GENI HOMEOBOX Motivo (homeodomain) conservato helix-turn-helix di binding al DNA di 180 bp
Classi di fattori di trascrizione
Mutazione knotted 1 (kn 1) identificata originariamente nel mais Mutazione gain of function (dominante) ØEspressione del gene nel momento e nel posto errato ØSi ha espressione di KN 1 nelle foglie durante lo sviluppo ØAnormalità intorno alle venature Proliferazione irregolare di divisioni cellulari intorno alle nervature Formazione di nodi (knot) che protrudono dalla foglia Nodi simili a meristemi, continuano a dividersi KN 1 controlla l’attività meristematica
Piante di tabacco trasformate con KN 1 sotto il controllo di un promotore costitutivo sviluppano meristemi apicali avventizi sulla superficie delle foglie KN 1 controlla l’attività dei meristemi
Nelle piante diverse sottofamiglie di geni HOMEOBOX Arabidopsis: ØKNOX (sottoclassi KNOXI , KNOX 2) ØWOX ØBELL ØHD-ZIP
In Arabidopsis geni di classe KNOX I KNAT 1 (BREVIPEDICELLUS) KNAT 2 KNAT 6 SHOOT MERISTEMLESS (STM) KNAT 1 e STM strettamente correlati
dimerizzazione repressione PEST loc nucleo DNA binding
In arabidopsis geni KNOX I espressi nelle diverse zone del SAM con pattern di espressione distinguibili ed in parte sovrapposti I geni KNOX I non vengono espressi nelle zone periferiche daranno luogo agli organi: per es nei primordi dei cotiledoni durante l’embriogenesi, o nei primordi fogliari nello sviluppo post-embrionale
I geni KNOX I sono essenziali per la formazione del SAM e per il suo mantenimento e organizzazione mutazione omozigote stm-1 meristema sulla superficie adaxiale di foglie di piante con espressione ectopica di STM
Mutanti SHOOTMERISTEMLESS (stm): STM espresso specificamente nelle cellule che diverranno cellule meristematiche Mutanti stm omozigoti, loss of function, non formano meristema apicale e le cellule si differenziano STM inibisce il processo di differenziamento assicurando che le cellule meristematiche rimangano indifferenziate STM è necessario anche per il mantenimento dell’identità meristematica delle cellule nella pianta adulta
STM L’espressione di STM richiede l’espressione dei geni CUC Successivamente STM restringe l’espressione dei geni CUC alla periferia della zona meristematica Durante la crescita vegetativa STM è espresso in tutto il meristema ma non nelle cellule fondatrici del tessuto fogliare e nei primordi fogliari
L’esclusione dei trascritti dei geni KNOX I dalle cellule iniziali e dai primordi ai lati del SAM implica che la funzione KNOX sia incompatibile con il differenziamento delle foglie. Funzione KNOX antagonistica al normale differenziamento ed espansione cellulare Meccanismi regolativi per escludere l’espressione di KNOX negli organi in differenziamento
ØNelle piante a differenza degli animali, l’organogenesi è postembrionale, consentendo di adattare lo sviluppo alle condizioni ambientali (piante organismi sessili) ØDopo la germinazione SAM dà luogo agli organi laterali ØSAM mantiene una struttura organizzata pur rispondendo a segnali interni ed esterni di sviluppo ØA questo scopo al centro del SAM viene mantenuta una popolazione di cellule indifferenziate che si dividono lentamente ØCellule che lasciano questa zona entrano in quelle periferiche e si differenziano a formare gli organi laterali; oppure nella zona sottostante (rib zone) e si differenziano in cellule del fusto La velocità di proliferazione delle cellule iniziali nel SAM deve essere coordinata con la velocità di differenziamento delle cellule figlie
Formazione e mantenimento del SAM: rete regolativa Implicati altri geni HOMEOBOX WUSCHEL
Il fenotipo stm è simile in apparenza a quello wus, in cui le piccole cellule in divisione del SAM perdono il loro potenziale meristematico determinando l’arresto dello sviluppo del SAM stm: cellule meristematiche consumate nello sviluppo degli organi wus: rimangono in uno stato indifferenziato non meristematico STM: richiesto per prevenire il differenziamento WUS: necessario per mantenere lo stato staminale
Nello sviluppo del SAM operano due pathways regolativi principali: ØIl circuito KNOX (STM) che regola il destino delle cellule meristematiche ØIl circuito WUS/CLV che regola l’ identità cellulare e il numero di cellule staminali nella zona centrale (CZ) del SAM
WUSCHEL (WUS) (gene homeobox di classe wox) espresso nel centro organizzatore (OC) sotto la zona centrale del SAM conferisce identità staminale alle cellule dello strato superiore Nei mutanti wus loss of function l’attività meristematica (SAM) cessa alla fine dell’embriogenesi e la crescita si arresta allo stadio di cotiledoni. Non viene mantenuta la popolazione delle cellule staminali
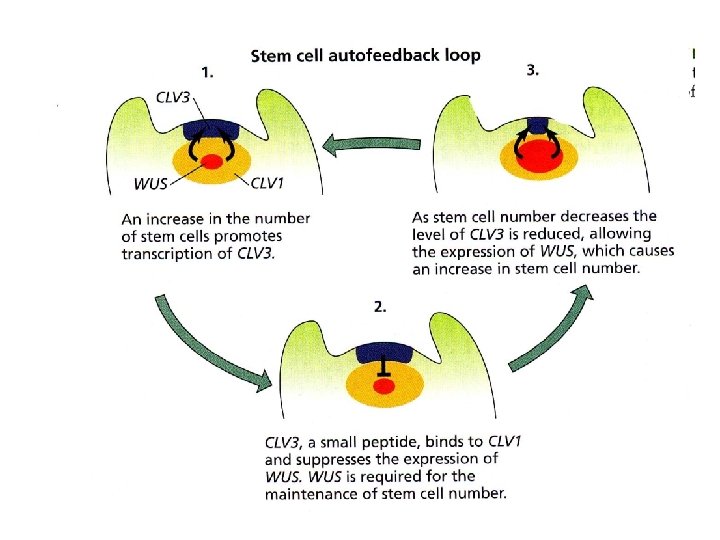
IDENTIFICATO UN CIRCUITO REGOLATIVO CHE COMPRENDE I GENI WUSCHEL (WUS) E CLAVATA (CLVI, CLV 2, CLV 3)
Il meccanismo di mantenimento del SAM e di coordinamento con il processo differenziativo identificato mediante lo studio dei mutanti di Arabidopsis WUSCHEL , CLAVATA-1, CLAVATA-2 , CLAVATA-3
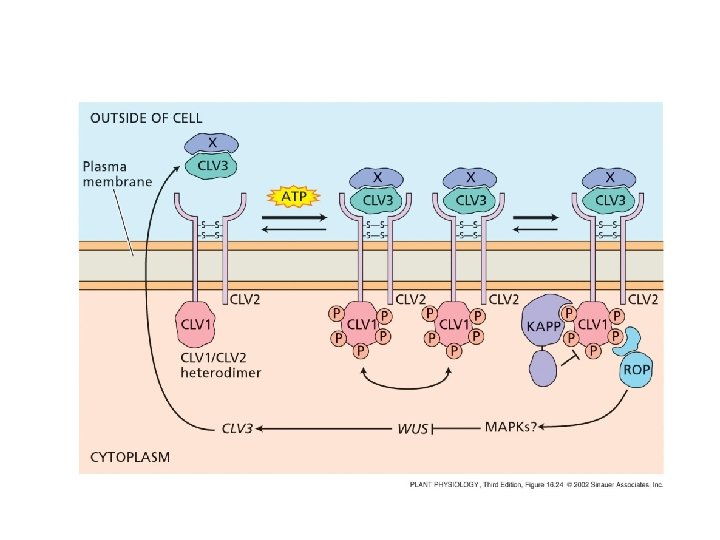
CLAVATA 1 è un recettore chinasi (receptor kinase) CLAVATA 2 manca del dominio chinasico ma possiede il dominio di binding per CLV 3 CLAVATA 3 è una piccola proteina appartenente alla classe CLE ed è il ligando di CLV 3/CLV 2
cascata di MAP chinasi
BRI 1 Recettore di sistemina e brassinosteroidi Xa 21 Clavata 1 Dominio Chinasico Serina/treonina kinasi
ØI geni CLAVATA sono stati identificati come mutazioni che determinano un aumento delle dimensioni dei meristemi vegetativo apicale e fiorale, con aumento nel numero degli organi laterali (numero di organi fiorali) ØI geni CLAVATA regolando l’espressione di WUSCHEL controllano le dimensioni del meristema apicale (numero di cellule iniziali nella zona centrale del SAM) MECCANISMO A FEEDBACK
WUS è espresso nelle cellule del centro organizzatore tra gli strati L 1 e L 3 nella zona centrale del SAM
CLV 3 è espresso nelle cellule degli strati L 1 e L 2 della CZ nel meristema apicale WUS è espresso nelle cellule del OC della zona centrale (strato L 3) CLV 1 ha un pattern di espressione simile a quello di WUS ma più ampio WUS agisce in maniera “non autonoma” infatti la sua attività è richiesta per mantenere l’identità delle cellule iniziali ma è espresso solo in poche cellule dello strato L 3 (azione a distanza) Se mutazioni inattivano CLV 1 o CLV 3 l’epressione di WUS si espande e aumenta il numero di cellule iniziali indifferenziate
Nei mutanti clv il doma ha dimensioni maggiori. Ciò è dovuto ad una maggiore estensione della ZC per il mancato differenziamento delle cellule nelle ZP In questi mutanti WUS ha un’espressione più ampia Piante transgeniche overesprimenti WUS hanno un fenotipo simile a clv Piante transgeniche overesprimenti CLV 3 hanno un fenotipo simile a wus Nel doppio mutante clv 3 wus il doma ha dimensioni normali
CLAVATA 3 regola negativamente l’espressione di WUSCHEL regola positivamente l’espressione di CLAVATA 3
STM mantiene la proliferazione cellulare nella zona periferica svolgendo un ruolo complementare a quello di WUSCHEL
STM e WUS agiscono in maniera complementare e indipendente ØL’espressione ectopica di STM nei primordi fogliari (ANT: : STM) sopprime il differenziamento. Si formano organi laterali aggiuntivi Si ha attivazione dei geni KNAT e repressione dell’espressione di CLV 3 (STM promuove l’attività proliferativa ma non l’identità di cellule iniziali) ØL’espressione ectopica di WUS (GR: : WUS) nei primordi fogliari induce l’espressione di geni di identità meristematica in un tessuto differenziato (CLV 3)
Espressione ectopica di ANT: : WUS in stm 5 background Espressione ectopica di ANT: : STM in wus 1 background fenotipi indistinguibili (o quasi) da wt background STM e WUS agiscono indipendentemente
STM sopprime il differenziamento indipendentemente da WUS ed è Sufficiente per promuovere la divisione cellulare WUS conferisce l’identità staminale indipendentemente da STM WUS e STM attivano geni diversi a valle I pathways regolati da WUS e STM sembrano convergere nella soppressione del differenziamento (l’espressione ectopica contemporanea di STM e WUS ha un effetto sinergico)
In conclusione…. STM e WUS si “dividono il lavoro” nel meristema apicale del germoglio WUS specifica una popolazione particolare di cellule nel centro del meristema (cellule staminali) STM serve a reprimere il differenziamento attraverso tutto il doma, consentendo alle cellule derivate di venire amplificate prima di essere incorporate negli organi in formazione
Le funzioni dei geni STM e WUS sono interconnesse a vie di signaling ormonali CITOCHININE GIBBERELLINE