Hearing Equilibrium The inner ear labyrinth is made

Hearing &Equilibrium
 is made up of two parts; one within the other,")
The inner ear (labyrinth) is made up of two parts; one within the other, the (bony labyrinth) is a series of channels in the petrous portion of the temporal bone. Inside these channels, surrounded by a fluid called (perilymph) is the (membrane labyrinth). This membrane structure more or less duplicates the shape of the bony channels. It is filled with a fluid called (endolymph), and there is no communication between the space filled with endolymph and those filled with perilymph.

Cochlea: The cochlear portion of the labyrinth is a coiled tube which in human is 3. 5 mm long and makes 2¼ turns. Throughout its length, the basilar membrane and Reissner`s membrane divided it into three chamber’s (scalae). The upper scala vestibuli and the lower scala tympani contain perilymph and communicate with each other at the apex of the cochlea through a small opening called the helicortrema. At the base of the cochlea, the scala vestibuli ends at the oval window, which is closed by the footplate of the stapes. The scala tympani end at the round window, a foramen on the medial wall of the middle ear that is closed by the flexible secondary tympanic membrane.
; Separate scala vestibuli and scala media")
A. Reissner`s membrane (also called the vestibular membrane); Separate scala vestibuli and scala media is so thin and so easily moved that it does not obstruct the passage of sound vibrations through fluid from the scala vestibuli into scala media. B. basilar membrane Separate scala media from scala tympani project from the bony center of the cochlea, the modiolus, toward the outer wall fibers are stiff, elastic, reed ﻧﺎﻱ ﺍﻭ ﻗﺼﺐ like structure contains 20, 000 to 30, 000 basilar fibers fixed at their basal ends in the central bony structure of the (the modiolus) but not fixed at their distal ends. Because the fibers are stiff and free at one end, they can vibrate like the reeds of a harmonica On the surface of the basilar membrane lies the

The basilar membrane characterized by: 1. The lengths of the basilar fibers increase progressively beginning at the oval window and going from the base of the cochlea to the apex, increasing from a length of about 0. 04 millimeter near the oval and round windows to 0. 5 millimeter at the tip of the cochlea (the “helicotrema”), a 12 -fold increase in length + 2. The diameters of the fibers, however, decrease from the oval window to the helicotrema so their overall the basilar fibers stiff ness decreases more than 100 -fold toward the helicotrema. ▼ high coef ficient of elasticity of the basilar fibers near the oval window Low coef ficient of elasticity of the basilar fibers near the helicotrema As a result, the short + stiff fibers near the oval window of the cochlea vibrate best at a very high frequency, Long + limber ﻣﺮﻥ ﺭﺷﻴﻖ fibers near the tip of the cochlea vibrate best at a low frequency. 3. The vibrating sound wave travels fast along the initial portion of the basilar membrane The vibrating sound wave travels slow near the helicotrema
: The initial effect of")
Transmission of sound waves in the cochlea (the traveling wave): The initial effect of a sound wave entering at the oval window is to cause the vibrating the vestibular membrane and this will cause basilar membrane bend in the direction of the round window. Therefore, when the foot of the stapes moves inward against the oval window the round window must bulge outward because the cochlea is bounded on all sides by bony walls. However, the elastic tension that is built up in the basilar fibers as they bend toward the round window initiates a fluid wave that travels along the basilar membrane toward the helicortrema. Each wave is relatively weak at the outset but becomes strong when it reaches that portion of the basilar membrane that has a natural

Consequently, the wave dies out at this point and fails to travel the remaining distance along the basilar membrane. Thus, a high-frequency sound wave travels only a short distance along the basilar membrane before it reaches its resonant point and dies out; a medium-frequency sound wave travels about halfway and then dies out; and finally, a very low frequency sound wave travels the entire distance along the membrane. This rapid initial transmission of the wave allows to spread out and separate from one another on the basilar membrane. Without this rapid initial transmission, all the high-frequency waves would be bunched ﺗﺘﺰﺍﺣﻢ together within the first millimeter or so of the basilar membrane, and their frequencies could not be discriminated. Organ of Corti: Located on the basilar membrane is the organ of Corti, the structure that contains the hair cells which the auditory receptors. This organ
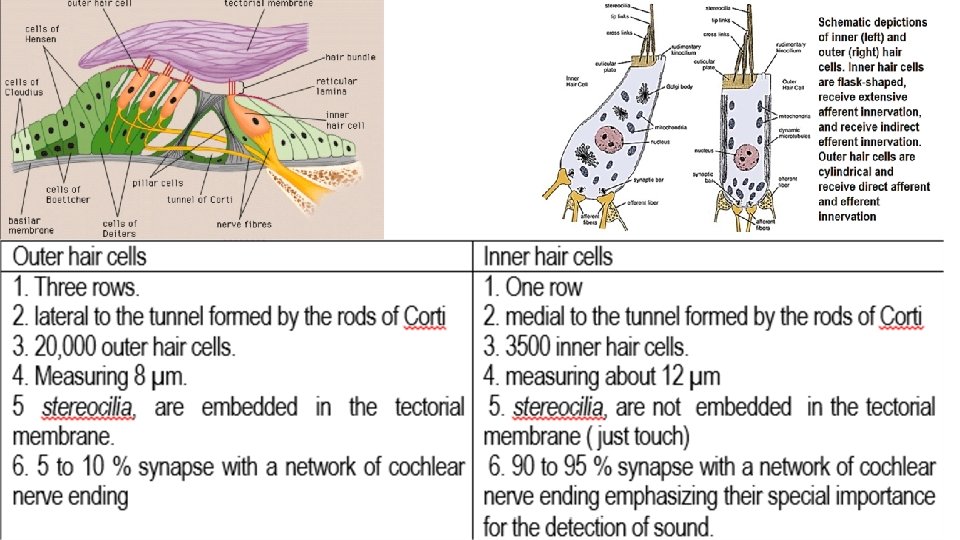

The cell body of the sensory neurons stimulated by the hair cells is located the spiral ganglion of Corti, which lies in the modiolus (center) of the cochlea. and then The spiral ganglion neuronal cells send axons (a total of about 30, 000) into the cochlear branch of vestibulo-chochlear Nerve (VIII nerve) into the central nervous system at the level of the upper medulla. The outer ends of the hair cells are fixed tightly in a rigid struc ture composed of a flat plate, called the reticular lamina, supported by triangular rods of Corti, which are attached tightly to the basilar fibers. The basilar fibers, the rods of Corti, and the reticular lamina move as a rigid unit.
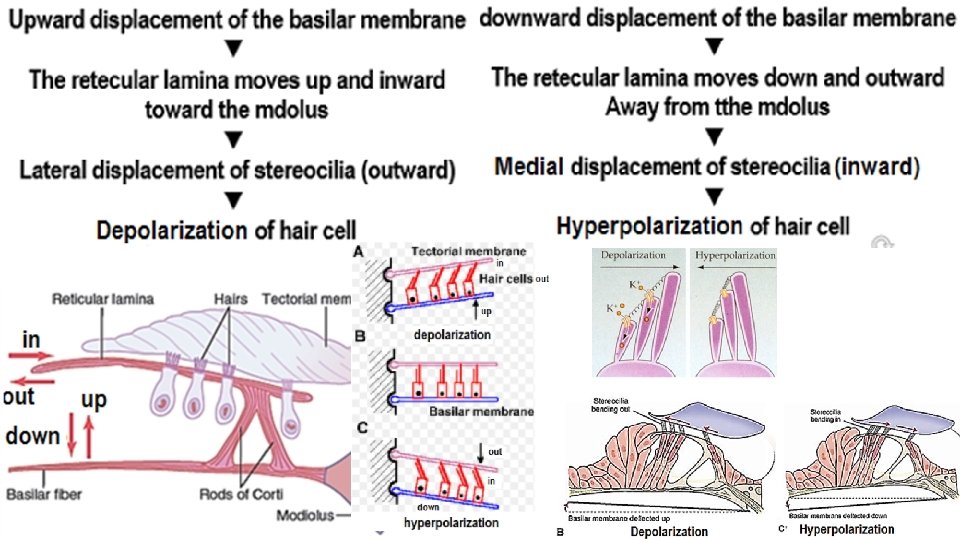

Depolarization of hair cell: Each hair cell has about 100 stereocilia on its apical border. These stereocilia become progressively longer on the side of the hair cell away from the modiolus When the hair bundle is displaced in the direction of the tallest stereocilium depolarization occurs When the hair bundle is displaced in direction away from the tallest stereocilium hyperpolarization occurs When the hair bundle is displaced in direction perpendicular to stereocilium provides no change in membrane potential When the hair bundle is displaced in direction intermediate between these two directions produces

The function of inner hair cells is transmission of hearing signals The function of outer hair cells If the outer cells are damaged while the inner cells remain fully functional, a large amount of hearing loss occurs. Outer hair cells contact by relatively few sensory axons and therefore accounted for only a small fraction of the auditory information that is sent onto the brain Unlike the inner hair cells, the outer hair cells do not signal the brain about incoming sounds. The principle function of outer hair cells is to amplify the sound-induce vibration so inner hair cell will respond more strongly and thus increase our ability to hear very quiet sounds

The outer hair cell do amplify sound by increasing and decreasing their length The outer hair cell actively and rapidly decreasing their length (shortening) occurs during depolarization and increasing their length (elongation) occurs during hyperpolarization. ▼ Amplify movement of tectorial membrane at area that matches the frequency of sound They do this due to: 1. a very flexible basal membrane, 2. prestin a trans-membrane protein that changes the length of outer hair cell. This behavior is known as electromotility. In support of this concept, a large number of retrograde nerve fibers pass from the brain stem to the vicinity ﺍﻟﻤﺠﺎﻭﺭﺓ ﺍﻟﻤﻨﺎﻃﻖ of the outer hair cells (i. e. this will explain why there is afferent and efferent nerve fiber to both outer and inner hair cells) ﺑﺘﻀﺨﻴﻢ ﺍﻟﺨﺎﺭﺟﻴﺔ ﻓﺘﻘﻮﻡ ﻭﺍﺿﺤﺔ ﻏﻴﺮ ﺍﻟﺪﺍﺧﻠﻴﺔ ﻣﻦ ﺍﺳﺘﻠﻤﺘﻬﺎ ﺍﻟﺘﻲ ﺍﻟﺸﺎﺭﺓ ﺍﻥ ﻋﻠﻰ ﺍﻟﺪﻣﺎﻍ ﻣﻦ ﺍﺷﺎﺭﺓ ﺗﺴﺘﻠﻢ ﺧﺎﺭﺟﺒﺔ

An electrical potential of about +80 millivolts exists all the time between endolymph and perilymph, with positiv ity inside the scala media and negativity outside. This is called the endocochlear potential, and it is generated by continual secretion of positive potassium ions into the
 Primary auditory cortex (AI; area")
• Function of cerebral cortex in hearing 1) Primary auditory cortex (AI; area 41) The primary auditory cortex is directly excited by auditory nerve The primary auditory cortex has tonotopic maps "frequency map" for different tones. The primary auditory cortex unilateral lesions do not effect hearing because of completely bilateral sound representation.

Sound frequency perception in the primary auditory cortex. Neurons in the auditory cortex are organized according to the frequency of sound to which they respond best. Neurons at one end of the auditory cortex respond best to low frequencies; neurons at the other respond best to high frequencies. The purpose of this frequency map (known as a tonotopic map) is unknown The frequency range to which each individual neuron in the auditory cortex responds is much narrower than that in the cochlear (i. e. “sharpen” the frequency). It is believed that this sharpening effect is caused mainly by the phenomenon of lateral inhibition. The same effect has been demonstrated to be important in sharpen ing patterns of somesthetic images, visual images, and other types of sensations.
 Auditory association cortex or the secondary auditory cortex (AII; area 42) The auditory")
2) Auditory association cortex or the secondary auditory cortex (AII; area 42) The auditory association areas surrounding the primary auditory cortex The auditory association areas are excited secondarily by impulses from the primary auditory cortex, as well as by some projections from thalamic association areas The auditory association areas are involved in the interpretation ﺗﺮﺟﻤﺔ of sound: Many of the neurons in the auditory cortex, especially in the auditory association cortex, respond to specific sound frequencies in the ear. “associate” different sound frequencies with one another associate sound information with information from other sensory areas of the cortex. Example association of audi tory information
 Wernicke's area (Brodmann’s area 22) Wernicke's area 22 is concerned with the processing")
3) Wernicke's area (Brodmann’s area 22) Wernicke's area 22 is concerned with the processing of auditory signals related to speech. Wernicke's area in the dominant hemisphere surrounding the auditory cortex Wernicke's area is much more active on the left side than on the right side during language processing Wernicke's area in the non-dominant hemisphere may be involved in understanding the tone ﻧﻐﻤﺔ pitch(frequency) sound intensity (loudness or amplitude) melody ﻟﺤﻦ The auditory pathways are also very plastic, and, like the visual and somasthetic pathways, they are modified by experience.

Discrimination of sound “Patterns” by the auditory Cortex. Destruction of one side primary auditory cortex a. only slightly reduces hearing in the opposite ear b. it does not cause deafness in the same ear because of many crossover connections from side to side in the auditory neural pathway c. it does affect one’s ability to localize the source of a sound, because comparative signals in both cortices are required for the localization function. Destruction of both primary auditory cortices in the human being greatly reduces one’s sensitivity for hearing. Lesions that affect the auditory association areas but not the primary auditory cortex do not decrease a per son’s ability to hear and differentiate sound tones, interpret at least simple patterns of sound. However, the person is often unable to interpret ﻓﺴﺮ the meaning of the sound heard even though he or she hears them perfectly well and can even repeat them ﺍﻻﺻﻮﺍﺕ ﻣﻌﺎﻧﻲ ﺗﻔﺴﺮ ﻋﻠﻰ ﺍﻟﻘﺪﺭﺓ ﻧﻔﻘﺪ ﻭﻟﻜﻦ ﺗﻜﺮﺍﺭﻫﺎ ﻋﻠﻰ ﺍﻟﻘﺪﺭﺓ ﻧﻔﻘﺪ ﻻ ﻭﺑﺎﻟﺘﺎﻟﻲ ﺍﻻﺻﻮﺍﺕ ﺍﻭﺗﻔﺮﻳﻖ ﺍﻟﺴﻤﺎﻉ ﻋﻠﻰ ﺍﻟﻘﺪﺭﺓ ﻧﻔﻘﺪ ﻻ Therefore, the auditory association areas cortex is especially important in

Determination of the direction from which sound come A person determines the horizontal direction from which sound comes by two principal means: (1) The intensity mechanism determine the distance depending on the difference between the inten sities of the sounds reaching the two ears. The intensity mechanism oper ates best at higher frequencies because the head is a greater sound barrier at these frequencies. If a person is looking straight toward the source of the sound, the sound reaches both ears at exactly the same instant If the right ear is closer to the sound than the left ear First, the paths are of different length because sound has to travel past the head to get to the left
 The time lag ﺗﺄﺨﺮ mechanism: The time lag between the entry of sound")
2) The time lag ﺗﺄﺨﺮ mechanism: The time lag between the entry of sound into one ear and its entry into the opposite ear (as little as 20 μs). This mechanism functions best at frequencies below 3000 cycles/sec The time lag mechanism discriminates direction much more exactly than the intensity mechanism because it does not depend on extraneous factors but only on the exact interval of time between two acoustical signals. The two aforementioned ﺍﻧﻔﺎ ﻣﺬﻛﻮﺭ mechanisms cannot tell whether the sound is emanating ﻣﻨﺒﺜﻖ from in front of or behind the person or from above or below. This discrimi nation is achieved mainly by the pinnae of the two ears. The shape of the pinna changes the quality of the sound entering the ear by empha sizing ﺗﺄﻜﻴﺪ specific sound frequencies depending on the
 the semicircular canals.")
Equilibrium The vestibular system has two parts, the otolith organs (macula) the semicircular canals. A mechanoreceptor (a hair cell with stereocilia) senses head position, head movement, and whether our bodies are in motion. The neural signals generated in the vestibular ganglion are transmitted through the vestibule-cochlear nerve to

1. Sensory organ of utricle and saccule; They are 2 millimeters in diameter They give information about static head position moving linearly the in 3 axis of the head The macula in the utricle and saccule contains an array of supporting cells and hair cells. The hair cells have 50 -70 stereocilia each and one large kinocilium. Hair cells stereocilia project into and embedded in the Otolithic membrane, contains a gelatinous mass otoliths (literally, “ear stones”) or otoconia (literally, “ear dust”). Otoconia are crystals of calcium carbonate range from 3 to 19 μm in length in humans Otoconia make the otolithic membrane heavier than the structures and fluids surrounding it.

Gravity pulls on the dense otoliths, which deform the gelatinous mass subsequently press on the stereocilia influence the firing rate of the hair cells. Since gravity pulls constantly, the hair cells in the utricle and saccule provide tonic information about the orientation of the head. The nerve fibers from the hair cells join those from the cristae in the vestibular division of the eighth cranial nerve
: 1. Linear acceleration and Head orientation")
A. Static vestibular apparatus First: (Macula of utricle): 1. Linear acceleration and Head orientation when person is upright position Utricle senses the Horizontal plane ﺍﻻﺭﺽ ﺳﻄﺢ ﻣﻮﺍﺯﻱ A. horizontal linear acceleration (motion in a straight line) , sense direction and acceleration The three axis linear motion X-axis (Moving forward and backward as in walking) ﺍﻟﺴﻴﺎﺭﺓ ﻓﻲ ﻛﻤﺎ Y-axis (Moving side to side) Z-axis (moving up and down as in elevator) B. Orientation of head when is upright Neck flexion dorsal and ventral Neck flexion right and left Vertebral Colum Angular movements of vertebral Colum (flexion and extension)

2. Macula of Saccule: The Macula of saccule senses the located in Vertical plane: ﻣﺴﺘﻮﻯ ﻣﻊ ﻋﻤﻮﺩﻱ ﺍﻻﺭﺽ Sense the head and body positions when the person is lying down For the Macula of saccule we can applied the same movement as we has discussed for Macula of utricle except instead of upright position lying down Note that the saccular and utricular maculae on one side of the head are mirror images of those on the other side. Thus, a tilt of the head to one side has opposite effects on corresponding hair cells of the two utricular maculae. This concept is important in understanding how the central connections of the vestibular periphery mediate the interaction of inputs from the two sides of the head When the stereocilia and kinocilium bend in the direction of the kinocilium causing recep tor membrane depolarization this will increase firing rated of nerve above 100 per second. The reverse will causes receptor hyperpolarization this will increase firing rated of nerve below 100 per second in same manner as explained above.

It is especially important that the hair cells are all oriented in different directions in the maculae of the utricles and saccules so that with different positions of the head, different hair cells become stimulated. This utricle and saccule system functions extremely effectively for maintaining equilibrium when the head is in the nearvertical position. Indeed, a person can determine as little as half a degree of disequilibrium when the body leans from the precise upright position. The “patterns” of stimulation of the different hair cells apprise ﺧﺒﺮ the brain of the position of the head with respect to the pull of gravity. In turn, the vestibular, cerebellar, and reticular motor nerve systems of the brain excite appropriate postural muscles

The impulses generated from these receptors are partly responsible for labyrinth righting reflexes. These reflexes are a series of responses integrated for the most part in the nuclei of the midbrain. The stimulus for the labyrinth righting reflexes is tilting of the head, which stimulates the otolithic organs; the response is compensatory contraction of the neck muscles to keep the head level. In cats, dogs, and primates ﻗﺮﻭﺩ , visual cues ﺍﻟﺒﺼﺮﻳﺔ ﺷﺎﺭﺍﺕ ﺃﻸ can initiate optical righting reflexes that right the animal in the absence of labyrinthine or body stimulation. In humans, the operation of these reflexes maintains the head in a stable position the eyes fixed on visual targets despite movements of the body By remarkably precise reflex contractions of the neck and extra-ocular muscles. The responses are initiated by vestibular stimulation, stretching of neck muscles, movement of visual images on the retina, and the responses are the vestibulo-ocular reflex

Although most of the responses to stimulation of the maculae are reflex in nature, vestibular impulses also reach the cerebral cortex. These impulses are presumably responsible for conscious perception of motion and supply part of the information necessary for orientation in space. Spatial orientation Orientation in space depends in part on input from the vestibular receptors visual cues ﺑﺼﺮﻳﺔ ﺩﻻﻻﺕ proprioceptors cutaneous extero-ceptors, especially touch and pressure receptors. These four inputs are synthesized at a cortical level into a continuous picture of the individual orientation in space. Vertigo is the sensation of rotation in the absence of actual rotation and is a prominent symptom when one labyrinth is inflamed.
 membranous labyrinth: Rotational movement; Semicircular canal: The three semicircular")
B. The Kinetic (crista ampullaris) membranous labyrinth: Rotational movement; Semicircular canal: The three semicircular ducts in each vestibular apparatus, known as the anterior, poste rior, and lateral (horizontal) semicircular ducts, are arranged at right angles to one another so that they rep resent all three planes in space. Each of the three semicircular canals senses just a one-dimensional component of rotational acceleration. The three semicircular canals give information about dynamic head position Note: head rotation in upright position is responsibility of semicircular canal at that dimension

Inside the bony canals, the membranous canals are suspended in perilymph. A receptor structure, the crista ampullaris, is located in the expanded end (ampulla) of each of the membranous canals. Each crista consists of hair cells and supporting (sustentacular) cells surmounted by a gelatinous partition (cupula) that closes off the ampulla. The processes of the hair cells are embedded in the cupula, and the bases of the hair cells are in close contact with the afferent fibers of the vestibular division of the eighth cranial nerve.

Responses to rotational acceleration Rotational acceleration in the plane of a given semicircular canal stimulates its crista. The response passes into the following stages: First: The endolymph, because of its inertia, is displaced in a direction opposite to the direction of rotation. The fluid pushes on the cupula, deforming it. This bends the processes of the hair cells. This will increase the number of nerve impulses (depolarization)

Second: When a constant speed of rotation is reached, the fluid spins at the same rate as the body and the cupula swings back into the upright position in the middle of the ampulla because of its own elastic recoil This will returns the number of nerve impulses back to basilar level gradually Third: When the rotation suddenly stops, exactly opposite effects take place: The endolymph continues to rotate while the semicircular duct stops. This time, the cupula bends in the opposite direction, causing the hair cell to stop discharging entirely (hyperpolarization)

Fourth: the endolymph stops moving and the cupula gradually returns to its resting position in 25 to 30 s, thus allowing hair cell discharge to return to its normal tonic level This will returns the number of nerve impulses back to basilar level gradually Movement of the cupula in one direction commonly causes an increase in the firing rate of single nerve fibers from the crista, whereas movement in the opposite direction commonly inhibits neural activity. Rotation causes maximal stimulation of the semicircular canals most nearly in the plane of rotation. Because the canals on one side of the head are a mirror image of those on the other side, the endolymph is displaced toward the ampulla on one side and away from it on the other. The pattern of stimulation reaching the brain therefore varies with the direction as well as the plane of rotation. Linear acceleration probably fails to displace the cupula and therefore does not stimulate the cristae. However, there is considerable evidence that when one part of the labyrinth is destroyed, other parts take over its functions.
- Slides: 34