Glukoneogenese Syntese av glukose Anabole reaksjonsveier Winnie Eskild

Glukoneogenese Syntese av glukose Anabole reaksjonsveier Winnie Eskild, IMBV 2004

Glukoneogenesen • Glukoneogenesen er en reaksjonsvei som produserer glukose ut fra enkle trekarbonatom metabolismeintermediater – Pyruvat, laktat, glyserol, -ketosyrer • Foregår i alle dyr, planter, sopp og mikroorganismer • I dyr er leveren hovedsete for glukoneogenese, noe foregår i nyrebark • Biomolekyler som kan omdannes til glukose kalles glukogene forbindelser • Alle sitronsyresyklusintermediater kan omdannes til glukose via oksaloacetat • De fleste aminosyrers karbonskjelett omdannes til glukose via SSS. Alanin (pyruvat) og glutamin/glutamat ( -ketoglutarat) er kvantitativt viktigere fordi disse finnes i høyere konsentrasjoner enn andre aminosyrer

Glukogene forbindelser

Glukoneogenesen • I motsetning til planter kan ikke dyr omdanne vanlige fettsyrer til glukose • Årsaken er at det ikke dannes noe nettoprodukt når acetyl-Co. A sendes inn i sitronsyresyklusen • Plantenes glyoksylatsyklus fører til nettodannelse av succinat som lett omdannes til glukose • Fettsyrenedbrytning gir viktige bidrag til glykoneogenesen i form av ATP og NADH

Glyoksylat syklus

Glukoneogenesen • Organismen er helt avhengig av tilgang på glukose fordi sentrale funksjoner i kroppen ikke klarer seg med andre energisubstrater • Organer som bruker glukose som hovedenergikilde: hjernen, nervesystemet, nyrene, testikler, røde blodceller, embryonale vev • Hjernen forbruker 120 g glukose pr dag
 til glukose • Prinsipielt samme reaksjonsvei i alle")
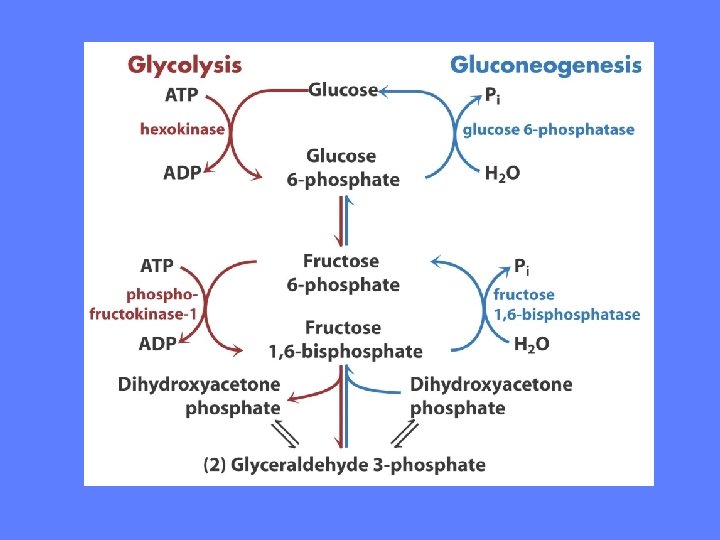
Glukoneogenesen • Omdannelse av karbon-forbindelser (pyruvat) til glukose • Prinsipielt samme reaksjonsvei i alle organismer og organer, små variasjoner i detaljene • Mens glykolysen omdanner glukose til fosfoenolpyruvat/pyruvat omdanner glukoneogenesen fosfoenolpyruvat til glukose

Glukose Pyruvat ATP ATP Glukose-6 -fosfat Oksaloacetat Fruktose-6 -fosfat GTP ATP NADH ATP Fosfoenol-pyruvat Fruktose-1, 6 -bisfosfat NADH ATP

Glykolyse vs glukoneogenese • A B • Motsatt rettede reaksjonsveier har oftest en del reaksjoner felles i tillegg til en eller flere som er unike for den enkelte reaksjonsveien • De unike reaksjonene er kontrollpunktene • Glukoneogenesen har 3 unike reaksjoner som atskiller seg fra glykolysen, resten av enzymene er felles • Unike enzymreaksjoner som kontrollerer hastigheten av en reaksjonsvei er irreversible under de aktuelle forholdene i cellen

Irreversible reaksjoner i glykolysen
 A) Heksokinase: 1) glc + ATP => glc-6")
Unike, irreversible reaksjoner Glykolyse Glukoneogenese 1) A) Heksokinase: 1) glc + ATP => glc-6 -P 2) Fosfofruktokinase 1: 1) fru-1 -P + ATP => fru-1, 6 bis. P 3) Pyruvat kinase: • fosfoenolpyruvat + ADP => pyruvat + ATP B) C) Glukose-6 -fosfatase: glc-6 -P => glc + Pi Fruktose 1, 6 -bisfosfatase: Fru-1, 6 -bis. P => fru-6 -P + Pi 2 mitokondrielle multienzym reaksjonsveier
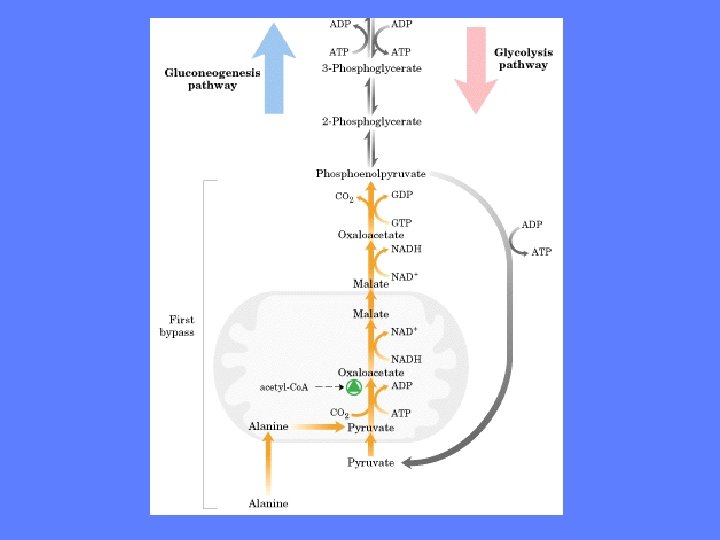

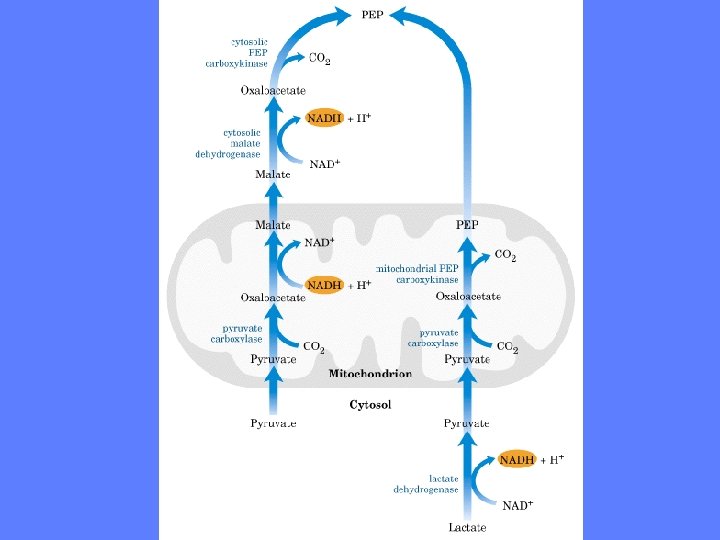
Glukoneogenesen - 1. unike reaksjonssekvens • Første reaksjonssekvens foregår i mitokondriet • I motsetning til glykolysen hvor fosfoenolpyruvat til pyruvat er én reaksjon trengs det en hel serie enzymreaksjoner til å omdanne pyruvat til fosfoenolpyruvat • Cellen har to forskjellige reaksjonsveier til å utføre omdannelsen av pyruvat til fosfoenolpyruvat. Substratet avgjør hvilken reaksjonsvei som velges. Mulighet for NADH produksjon er viktig • NADH trengs til å redusere 1, 3 -bisfosfoglyserat til glyseraldehyd-3 -P • Når alanin eller pyruvat er substrater velges den ene reaksjonsveien – Denne medfører at 2 elektroner i form av NADH ”hentes” ut fra mitokondriet • Når laktat er substrat velges den andre reaksjonsveien – Denne medfører at NADH dannes før pyruvat går inn i mitokondriet

Pyruvat til fosfoenolpyruvat - I • Pyruvat transporteres inn i mitokondriet eller dannes ved transaminering av alanin inne i mitokondriet • Pyruvat karboksylase katalyserer flg. reaksjon: Pyruvat + HCO 3 - + ATP => oksaloacetat + ADP + Pi. Biotin er kofaktor her • Dette er et viktig reguleringspunkt i glukoneogenesen. Pyruvat karboksylase reguleres positivt av acetyl-Co. A • Anaplerotisk reaksjon

Biotins reaksjonsmekanisme • Ureidogruppen bærer CO 2 • Enzymet har 2 aktive seter: Aktivering av bikarbonat Overføring av CO 2 til pyruvat

Pyruvat til fosfoenolpyruvat - I • • Þ • • Oksaloacetat reduseres til malat Enzymet er malat dehydrogenase (mitokondriell) Reduserende ekvivalenter leveres av NADH => Oksaloacetat + NADH + H+ <=> malat + NAD+ G = 0 k. J/mol. Reaksjonen er reversibel og inngår både i sitronsyresyklus og glukoneoenesen Malat overføres til cytosol via malat- ketoglutarat transportøren I cytosol reoksideres malat til oksaloacetat Malat + NAD+ => oksaloacetat + NADH + H+ Enzymet er den cytosoliske isoformen av malat dehydrogenase Resultatet av disse to reaksjonene er overføring av to elektroner til cytosol

Pyruvat til fosfoenolpyruvat - I • I cytosol reoksideres malat til oksaloacetat Þ Malat + NAD+ => oksaloacetat + NADH + H+ • Enzymet er den cytosoliske isoformen • av malat dehydrogenase • Resultatet av disse to reaksjonene er • overføring av to elektroner til cytosol

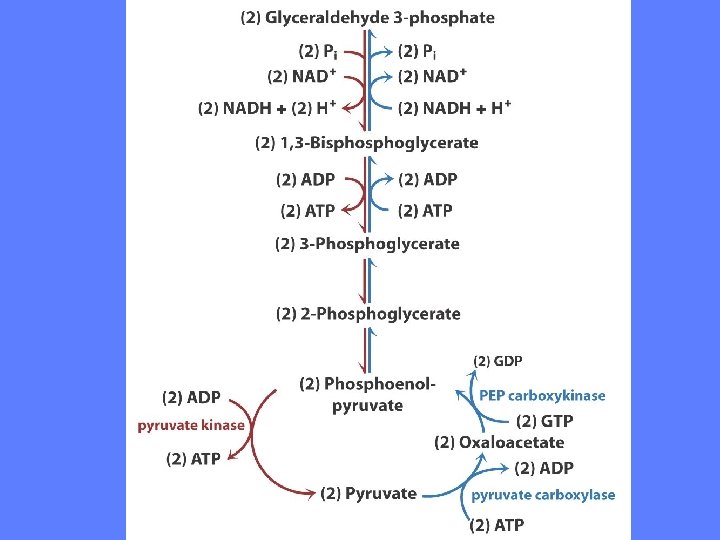
Pyruvat til fosfoenolpyruvat -I • Oksaloacetat dekarboksyleres og fosforyleres og blir til fosfoenolpyruvat • Enzymet er fosfoenolpyruvat karboksykinase Þ Oksaloacetat + GTP Mg 2+ fosfoenolpyruvat + GDP + CO 2 • Det avspaltede CO 2 er det samme som ble koplet på av pyruvat karboksylase • Reaksjonssekvensen kan ses som eksempel på aktivering ved karboksylering av et molekyle som skal fosforyleres (finnes også i fettsyresyntese)

Pyruvat til fosfoenolpyruvat - I • Nettoreaksjon: pyruvat + ATP + GTP + HCO 3 • fosfoenolpyruvat + CO 2 + GDP + ADP + Pi • G’o = 0, 9 k. J/mol • Konsentrasjonen av fosfoenolpyruvat holdes svært lav i cellen • Derfor er G = -25 k. J/mol • Overføring av elektroner til cytosol er essentielt for glukoneogenesen. NADH / NAD+ i cytosol er vanligvis 10 -4, mens det er ca 10 i mitokondriet

Pyruvat til fosfoenolpyruvat - II • • Þ • Når laktat er substrat går omdannelsen til fosfoenolpyruvat via en alternativ rute Laktat er resultat av anaerob glykolyse f. eks ved muskelarbeid eller normal energi-omsetning i røde blodceller Første reaksjon foregår i cytosol. Her oksideres laktat til pyruvat og de frigjorte elektronene overføres til NAD+ Nå er reduserende ekvivalenter for glukoneogenesen sikret Laktat + NAD+ pyruvat + NADH + H+ Enzymet er laktat dehydrogenase

Pyruvat til fosfoenolpyruvat - II • Pyruvat overføres til mitokondriet • Pyruvat omdannes til oksaloacetat i en karboksyleringsreaksjon katalysert av pyruvat karboksykinase Þ Pyruvat + ATP + HCO 3 - Mg 2+ oksaloacetat + ADP + Pi • Oksaloacetat kan ikke passere indre mitokondriemembran. Veien om malat er unødvendig siden NADH allerede er dannet i cytosol • Oksaloacetat omdannes direkte til fosfoenol-pyruvat av det mitokondielle isoenzymet fosfoenolpyruvat karboksykinase Þ Oksaloacetat +GTP Mg 2+ fosfoenolpyruvat + CO 2 + GDP • Fosfoenolpyruvat overføres til cytosol

Fosfoenolpyruvat til fruktose 1, 6 -bisfosfat • Serien av reaksjoner som omdanner fosfoenolpyruvat til fruktose 1, 6 -bisfosfat er alle reverserte glykolysereaksjoner Pi + aldolase ADP ATP

Glukoneogenesen - 2. unike reaksjon • Glykolysens fosfofruktokinase 1 reaksjon er irreversibel • I glukoneogenesen er det fruktose 1, 6 -bisfosfatase som hydrolyser bindingen til fosfatgruppen på C-atom 1 • G’o = - 16, 3 k. J/mol

Glukoneogenesens 3. unike reaksjon • Glykolysens heksokinasereaksjon er irreversibel • I glukoneogenesen er det enzymet glukose-6 -fosfatase som katalyserer hydrolyse av bindingen til fosfatgruppen. Enzymet er Mg 2+ avhengig • G’o = -13, 8 k. J/mol • Dette enzymet finnes bare i lever og nyre hvor det er assosiert med endoplasmatisk retikulum. Bare disse vev produserer glukose som sendes via blodsirkulasjonen til andre vev. Glukose -6 -fosfat kan ikke passere cellemebranen • Glukose-6 -fosfatase er assosiert med endoplasmatisk retikulum

Hvert glukosemolekyle koster 10 -12 ATP • G = -16 k. J/mol • Omkostning: 4 ATP + 2 GTP (=2 ATP) + 2 NADH (= 5 ATP) = 11 ATP

Pentosefosfat reaksjonsveien Syntese av NADPH og ribose-5 -fosfat Winnie Eskild, IMBV 2004

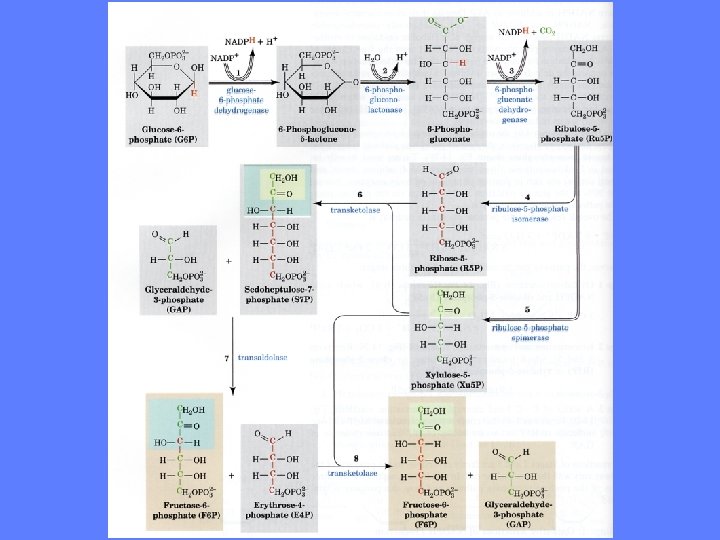
Pentosefosfatveien • Formålet med denne reaksjonsveien er at danne ribose-5 -fosfat og/eller NADPH • Pentosen ribose er nødvendig i syntesen av nukleinsyrer • NADPH er elektrondonor i de fleste synteseveier • Pentosefosfatveien er spesielt viktig i celler med aktiv steroid- og fettsyresyntese (f. eks. binyrer og fettvev) • I andre vev aktiveres pentosefosfatveien bare i forbindelse med celledeling hvor det må syntetiseres mye nukleinsyrer • Pentosefosfatveien deles i en oksidativ del og en non-oksidativ del – Oksidativ del: dannelse av ribose-5 -fosfat og NADPH – Non-oksidativ del: omdannelse av ribose-5 -fosfat til glukose-6 -fosfat

Dehydrogenering av glukose-6 -fosfat • C-1 i glukose-6 -fosfat dehydrogeneres og omdannes herved til en intramolekylær ester, 6 -fosfo-glukonod-lakton • Enzymet er glukose-6 -fosfat dehydrogenase som trenger Mg 2+ • Det ekstraheres 2 elektroner og 2 protoner fra glukose-6 -fosfat • 2 elektroner og 1 proton overføres til NADP+ som omdannes til NADPH • En proton frigis til miljøet

Hydrolyse av 6 -fosfoglukono-d-lakton • 6 -fosfoglukono-d-lakton hydrolyseres rask til 6 -fosfoglukonsyre som dissosierer til 6 -fosfoglukonat • Enzymet er laktonase og trenger Mg 2+ for full aktivitet

Dehydrogenering og dekarboksylering • 6 -fosfoglukonat dehydrogeneres og dekarboksyleres og omdannes herved til pentosen D-ribulose-5 fosfat • Enzymet er 6 -fosfoglukonat dehydrogenase som trenger Mg 2+ • Det overføres en hydridion (2 elektroner og en proton) til NADP+ og en proton frigis til miljøet • Dekarboksyleringen frigir CO 2 til miljøet

Isomerisering av ribulose-5 -fosfat • Ketosen ribulose-5 -fosfat isomeriseres til aldosen ribose-5 fosfat • Enzymet er fosfopentose isomerase

Oppsummering av de fire reaksjonene • Glukose-6 -fosfat + 2 NADP+ + H 2 O => ribose-5 -fosfat + 2 NADPH +2 H+ + CO 2 • Nettoutbytte er 2 NADPH til generelle synteseformål og et molekyle ribose-5 -fosfat til nukleinsyresyntese

Omdannelse av ribose-5 -fosfat til glukose 6 -fosfat • I vev som bare trenger NADPH, men ikke ribose-5 -fosfat, kan denne omdannes til glukose-6 -fosfat og igjen brukes til NADPH dannelse • Omdannelse av ribose-5 -fosfat til glukose-6 -fosfat er den nonoksidative delen av pentosefosfatveien • To enzymer er aktive her: – Transketolase som bruker tiaminpyrofosfat (TPP) som kofaktor – Transenolase

Epimerasereaksjonen • Ribose-5 -fosfat epimeriseres til xylulose-5 -fosfat • Epimerer har forskjellig konfigurasjon ved ett C-atom • Enzymene er epimerase og transketolase

Transketolasereaksjonene • Transketolaser katalyserer overføringen av en -karbonatomenhet fra en ketose til en aldose 2

Transaldolasereaksjonen • Transaldolase overfører en 3 -karbonenhet fra en ketosedonor til en aldose mottaker

Transketolase overfører 2 -karbonenhet fra xylulose-5 fosfat til ribose-5 -fosfat Herved dannes sedoheptulose-7 -fosfat og glyseraldehyd-3 -fosfat

Transaldolasereaksjonen • Her overføres en 3 -karbonenhet fra sedoheptulose-7 -fosfat til glyseraldehyd-3 -fosfat • Vi får dannet fruktose-6 -fosfat og erytrose-4 -fosfat • Enzymet er transaldolase som overfører 3 -karbonenheter

Transketolasereaksjon • Det overføres en 2 C-enhet fra xylulose-5 -fosfat til erytrose-5 fosfat • Herved dannes fruktose-6 -fosfat og glyseraldehyd-3 -fosfat • Enzymet er transketolase

Omdannelse til glukose-6 -fosfat • De to fruktose-6 -fosfat isomeriseres til glukose-6 -fosfat • Enzymet er fosfoheksoseisomerase • To glyseraldehyd-3 -fosfat omdannes til glukose-6 -fosfat via fruktose-6 -fosfat • Enzymene triosefosfat-isomerase, aldolase, fruktose-1, 6 -bisfosfatase • Totalt har vi fått omdannes 6 ribose-5 -fosfat til 5 glukose-6 fosfat

Ikke-oksidativ del av pentosefosfatveien

Seks ribose-5 -fosfat omdannes til fem glukose-6 -fosfat
- Slides: 46