Generalidades de Nutricin y Metabolismo de los Protozoarios

Generalidades de Nutrición y Metabolismo de los Protozoarios Parásitos

Nutrición • Dependencia metabólica del hospedero • Capacidad biosintética limitada • Conocimiento derivado de cultivos y en pocas especies • Single omission tests: difícil sacar conclusiones • Sustancias nutritivas universales: HC, aa, vitaminas, minerales y oligoelementos. Se suman: nucleósidos, AG, esteroles y porfirinas

Nutrición - Mecanismos • Difusión simple: moléculas apolares, liposolubles, AG no disociados, drogas hidrofóbicas • Transporte mediado: PM bajo, iones, H, Na, Cl, aa, azúcares. No utiliza energía, cinética de saturación y especificidad • Transporte activo: sistemas de transporte contra gradiente de concentración • Endocitosis: pinocitosis y fagocitosis
 glucosa")
Glúcidos • Transporte mediado en membranas celulares • T. brucei: 2 sistemas, a) glucosa y manosa y b) fructosa y glucosamina • T. equiperdium: 1 sitio de hexosas y 1 de glicerol • E. histolytica: glucosa por 2 sitios, paso limitante • Plasmodium: modulación de la membrana de la célula huésped (eritrocito) con poros.

Caso: Metabolismo de Galactosa en T. brucei. • El metabolismo de la Gal es esencial para la sobrevida de T. brucei. • Gal está presente en cantidades importantes en las VSG • Los transportadores de hexosas de T. brucei son incapaces de transportar Gal que se obtiene por la epimerización de la UDP-glucosa a la UDP-galactosa por la UDPglucosa 4´epimerasa (gal. E).
")
Generación de mutante condicional para gal. E • Línea celular knock-out condicional (c. KO) conteniendo una copia ectópica de gal. E inducible por Tetraciclina y ambas copias alélicas del gen reemplazadas con genes resistentes al antibiótico. • En presencia de Tetraciclina (+Tet) las células c. KO expresan suficiente gal. E para crecer normalmente. • En (–Tet) se detiene la división en 3 -4 días seguida de muerte celular.

Efecto de la deprivación de Gal en el crecimiento de T. brucei y contenido de nucleótidos (A) Growth of gal. E-c. KO with and without Tet (B) ratio of UDP-Gal/UDP-Glc in the gal. E-c. KO Tet (C) Sugar nucleotide levels of the gal. Ec. KO Tet cells. • Los niveles celulares de UDP-galactosa caen rápidamente luego de la deprivación de Gal llegando a niveles indetectables a las 72 hs.

Impacto de la deprivación de Gal sobre las VSG El análisis de las glicoproteínas extraídas por lectin blotting muestra que la Gal está virtualmente ausente y que se reducen las estructuras de poli-N-acetillactosamina

Mutante condicional null para gal. E • Bajo condiciones no permisivas que induce la deprivación de Gal. • Por medio de la adición de Tetraciclina al medio se produce la expresión de gal. E • Después de 96 h la división cesa y la ME revela una morfología alterada y aparición de vesículas aberrantes cerca del bolsillo flagelar. Scanning electron microscopy of gal. E-c. KO Tet. Effects on cellular morphology after galactose starvation for 0 h (A), 48 h (B), 96 h (C and D), and 144 h (E and F) are shown. Scale bars, 2 m.

Impacto de la deprivación de Gal sobre las VSG El análisis por MALDI-TOF de una VSG (221) confirma la pérdida completa de galactosa del ancla de glicosilfosfatidylinositol

Aminoácidos • • Translocación – mediada en su gran mayoría Endocitosis de proteínas. T. brucei: 4 sitios operativos T. cruzi: Arg altamente específico, con 3 sistemas; Thr es contra gradiente, se intercambia con Ala • Plasmodium: Hb es la mayor fuente de aa, ingresa por endocitosis a través del citostoma

Caso: la Arginina es un aa esencial para Toxoplasma gondii • En la mayoría de los eucariotes existen dos genes de carbamoil fosfatasa sintetasa (CPS), una es glutamina dependiente y se requiere para sintetizar pirimidinas (CPSII), la otra (CPSI) está dedicada a la biosíntesis de Arg a partir de carbamoil fosfato • T. gondii carece del gen de la CPSI • Deprivación de Arg bloquea multiplicación en taquizoítos, lo rescata la citrulina.

Síntesis de la Arginina

Deprivación de Arg dispara transformación de taquizoítos en bradizoítos Verde: proteína expresada por taquizoítos Rojo: proteína expresada por bradizoítos Azul: núcleos teñidos con DAPI A: 48 hs medio normal B: 48 hs medio sin Arg C: 7 d sin Arg D: 14 d sin Arg E: Contraste de fase de D F: Cepa PLK 4 d sin Arg

La deprivación de Arg produce una detención del crecimiento en taquizoítos de T. gondii que es rescatada por citrulina pero no por ornitina

Caso: Degradación de la Hb en Plasmodium

Hemoglobina · 95% de las proteínas totales de GR · Abundante (>300 mg/ml o aprox 5 m. M) · 60 -80% es degradada durante el estadio eritrocitario · 110 g (en un total de 750) se consume en 48 hs con una parasitemia del 20%

Erythrocytic Shizogonic Cycle • Los merozoítos interactúan con la membrana del GR e invaden activamente la célula formando una vaculoa parasitófora Merozoíto entrando un GR Trofozoíto en GR
")
Endocitosis del citoplasma eritrocitario cytostome food vacuole pinocytosis (rings)

Vacuola Digestiva Un lisosoma especializado Digestión de Hb Camino endocítico ATP H+ (p. H 5 -5. 4) Proteases de la vacuola • plasmepsinas I & II (acid) • falcipainas I - III (tiol) • falcilysinas (metallo) ADP citoplasma parasitario

El clivaje inicial de las plasmepsina es específico y desestabiliza la Hb • Hb es clivada entre Phe-33 y Leu -34 (cadena α) – ‘región bisagra’ – conservada – Importante para la estabilidad del tetrámero a-F 33/L 34 í • Se disocian grandes fragmentos de globina – Se libera el grupo Hem • Los fragmentos de globina son susceptibles de posterior hidrólisis

La digestion de Hb es un proceso ordenado hemoglobina plasmepsina fragmentos + heme grandes de globinas falcipaina plasmepsina Fragmentos medianos (20 aa) falcilysina • exopeptidasas • aa libres? Fragmentos pequeños (6 -8 aa)

El Heme libre es tóxico • Destabiliza and lisa membranas • Hidrolasas liberadas al citoplasma del parásito que muere Posibles Mecanismos de Detoxificación heme hemozoína (pigmento malárico) • Degradación mediada por H 2 O 2 • Degradación mediada por GSH • heme oxygenasa (P. b. and P. k. )

Hemozoína = b-Hematina heme b-hematin

b-hematina forma cristales insolubles 'biocristalización' o 'biomineralización'
 de novo • Purinas")
Nucleótidos • No pueden sintetizar nucleótidos de purinas (A G) de novo • Purinas y pirimidinas deben adquirirse en forma de bases o nucleósidos. No tienen transportadores de nucleótidos • En kinetoplástidos Adenosina es fuente más importante de síntesis de nucleótidos de purina • Locus para transporte de nucleósidos • Ribonucleasas y nucleotidasas de superficie • Plasmodium: hipoxantina es fuente de purina

Vías de salvataje de purinas en parásitos

Lípidos • Transportadores para AG • Difusión pasiva para AG no disociados • Apicomplexa: transporte a través de vacuola parasitófora • Apicoplasto: plástido no fotosintético encontrado en apicomplexa adquirido por endosimbiosis secundaria • Síntesis de AG e isoprenoides en apicoplasto

Metabolismo del colesterol en Toxoplasma gondii
 Catabolizar sustancias")
METABOLISMO ENERGÉTICO EN PROTOZOARIOS PARÁSITOS Generalidades Objetivos del metabolismo energético • a) Catabolizar sustancias orgánicas y acoplar el proceso a la conservación de energía • b) Formar y degradar biomoléculas requeridas en funciones específicas
 Depende únicamente de glicólisis para producir")
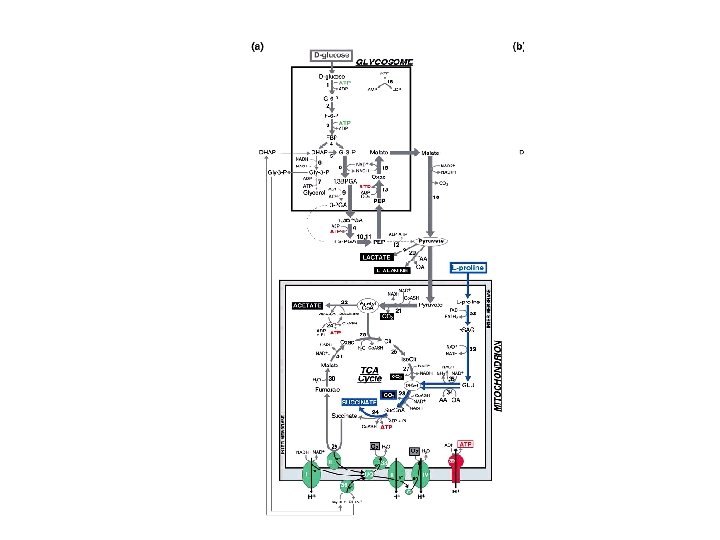
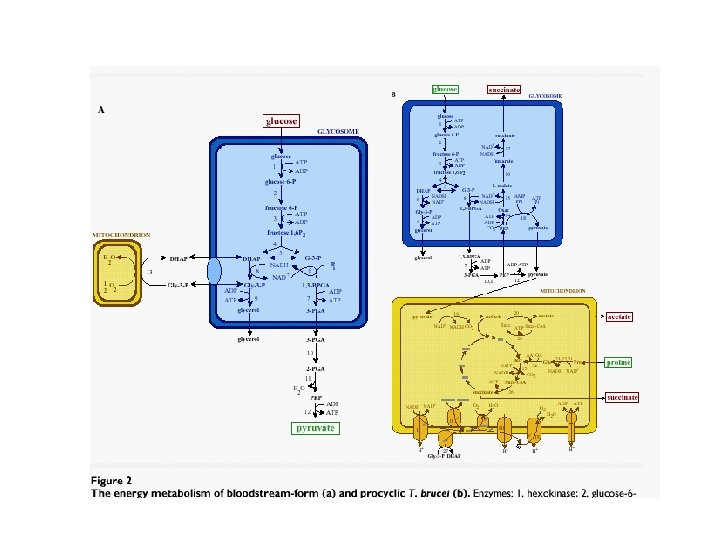
KINETOPLASTIDOS • Trypanosoma brucei como modelo: • a) Depende únicamente de glicólisis para producir ATP. Prefiere Glu, pero también Fru, Man y Glicerol • b) Mitocondria escasamente desarrollada sin Krebs ni CR • c) Abundantes enzimas glicolíticas: 90% del glicosoma • d) Flujo glicolítico es relativamente alto • e) Enzimas glicosómicas no glicolíticas deprimidas

Glicólisis • Glicosomas: Característicos de Tripanosomatidos. 0. 3 um, 4% del volumen celular. T. brucei=200. • Glicólisis: De Glu a 3 fosfoglicerato en glicosomas. Citoplasma: de 3 GP a 3 PEP – Piruvato. Termina en Piruvato (98%) y trazas de CO 2 y Glicerol- Fermentación G 3 P pasa los equivalentes reductores a través de una oxidasa al O 2 dentro de la mitocondria • Alta eficiencia. Sobrevive aún en condiciones anaeróbicas a razón de 1 ATP por 1 Glucosa. • Enzimas: sectores de carga + para ingresar al glicosoma. • Formas procíclicas en vector: Cambio a metabolismo más mitocondrial, aumenta volumen mitocondrial, cristas desarrolladas. CR respiratoria convencional. PRO: metabolizada en CO 2, ALA y ASP

Glicólisis y glicosomas •

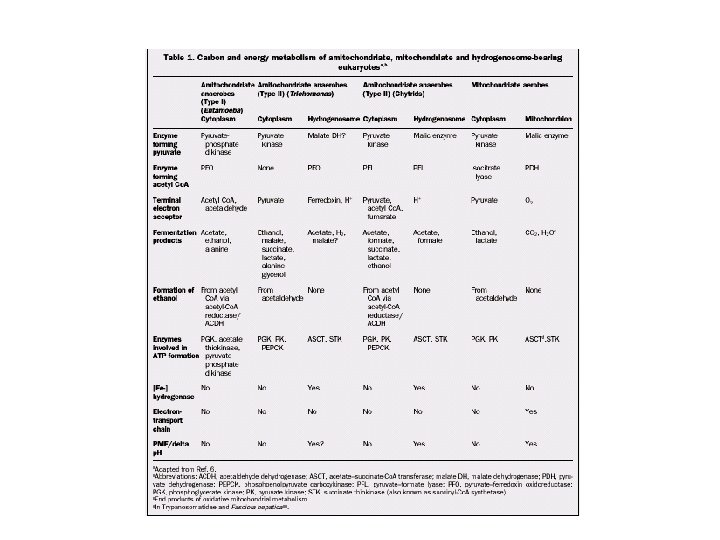
Amebas Intestinales y Giardia • Glicólisis via Embden-Meyerhof pero sin lactato deshidrogenasa. Piruvato se convierte en etanol y CO 2 en anaerobiosis, en aerobiosis acetato y etanol • Amitocondriados, por tanto sin Ciclo de Krebs ni fosforilación oxidativa • Sin citocromos • Almacenan glucógeno • Toleran bajas concentraciones de oxígeno

Figure 2 Nucleoside phosphate and pyrophosphate metabolism in the glycolytic pathway of the eukaryotic anaerobic amoeba, Entamoeba histolytica. (A) The glycolytic pathway of E. histolytica. (B) PPDK uses PEP and PPi to catalyze the conversion of AMP to ATP (i. e. , the production of two high-energy phosphoanhydride bonds equals two "ATP equivalents"). (C) The ATP and PPi "balance sheet" for E. histolytica glycolysis. Abbreviations are as follows: 1, 3 -DPGA, 1, 3 -diphosphoglycerate; Fru-1, 6 -P 2, fructose-1, 6 -bisphosphate; Fru-6 -P, fructose-6 -phosphate; Glu-6 -P; glucose-6 phosphate; 3 -PGA, 3 -phosphoglycerate; PEP, phosphoenolpyruvate; PPDK, pyruvate orthophosphate dikinase; and PPi. PFK, PPi-dependent phosphofructokinase

Enzimas glicolíticas de Entamoeba histolytica

Trichomonas • Glicólisis clásica hasta piruvato que se convierte en lactato y éste en acetato, CO 2 y H 2 O • La oxidación del piruvato se cataliza por decarboxilación oxidatica en reacciones ligadas a Ferredoxina, proteina sulfurada con Fe como transportador de electrones • La reacción se produce en los hidrogenosomas donde el H+ es el aceptor final de los electrones

Hidrogenosoma • Generación de ATP por compartimentalización del metabolismo fermentativo del piruvato con producción de Hidrógeno molecular • 1. Piruvato (de la glicólisis o por conversión del malato) es descarboxilado por la piruvato: ferredoxina oxidorreductasa para formar Acetil. Co. A • 2. Los e- son transferidos desde la oxidorreductasa hasta la ferredoxina (Fe-S) y luego a protones para formar H 2 catalizdo por la hidrogenasa. • 3. Acetil-Co. A es convertida a acetato con la conversión concomitante de succinato a Succinil-Co. A por la succinil. Co. A sintetasa. • 4. La generación de succinyl-Co. A está acoplada a la producción de ATP via fosforilación a nivel de substrato
![CO 2 hsp 70 ME Pyruvate Malate Transit peptides NAD(P)H NAD(P)+ [Fe]Hyd ATP ADP](http://slidetodoc.com/presentation_image_h/d3f1bac27d186e7e2263c7ad15e631b7/image-41.jpg "CO 2 hsp 70 ME Pyruvate Malate Transit peptides NAD(P)H NAD(P)+ [Fe]Hyd ATP ADP")
CO 2 hsp 70 ME Pyruvate Malate Transit peptides NAD(P)H NAD(P)+ [Fe]Hyd ATP ADP H 2 2 Fd- 2 H+ Co. ASH ASCT Acetate Acetyl-Co. A PFO CO 2 Fungi and Trichomonas Enzyme found also in mitochondria Alpha-proteobacterial ancestry Unknown ancestry AAC N NAD(P)-FO 2 Fd cpn 60 Protein import Succinate Succinyl-Co. A STK ATP Double membrane ADP + Pi Schematic Map of Hydrogenosomes (after Muller 1993)
- Slides: 42