Gene Regulation in Prokaryotes Outline of Chapter 16




























 n Physically")
















 of")
- Slides: 48
Gene Regulation in Prokaryotes
Outline of Chapter 16 n n There are many steps in gene expression and regulation can occur at any one of them Genetic and molecular studies show that most regulation affects the initiation of RNA transcripts n Studies of genes for lactose utilization n n Negative regulation – blocks transcription Positive regulation – increases transcription DNA binding proteins acting on RNA polymerase at promoter are main agents of regulation Attenuation of expression – tryptophan pathway n Gene expression is fine tuned by premature termination of transcription
Outline of Chapter 16 n Global regulatory mechanisms: E. coli’s response to heat shock is an example of the bacterial ability to coordinate the expression of different sets of genes dispersed around the chromosome. n n Microarray analysis is an important new tool for detecting changes in gene expression in response to environmental changes Comprehensive example: How Vibrio cholerae regulate their virulence genes
RNA polymerase is the key enzyme for transcription n RNA polymerase involved in three phases of transcription n Initiation – sigma subunit + core enzyme (two alpha, one beta, and one beta’ subunit) n n n Binds to promoter, unwinds DNA, begins polymerization of bases complementary to DNA template Elongation – movement away from promoter sigma subunit released, polymerization Termination –signal reached by RNA polymerase n n Rho dependent termination – Rho factor recognizes sequence in m. RNA, binds to it, and pulls it away from RNA polymerase Rho independent termination – stem loop structure formed by sequence of 20 bases with a run of 6 or more U’s signals release of RNA polymerase
Fig. 16. 2
n Translation in prokaryotes starts before transcription ends n n n Initiation sites for translation signal ribosomes to bind near 5’ end of m. RNA while downstream transcription is still occurring Polycistronic m. RNAs often lead to the translation of several genes at the same time from one m. RNA transcript The regulation of gene expression can occur at many steps n n n Binding of RNA polymerase to promoter Shift from initiation to elongation Release of m. RNA at termination Posttranscriptional stability of m. RNA Efficiency of ribosomes to recognize translation initiation sites Stability of polypeptide product
The utilization of lactose by E. coli: A model system for gene regulation n The presence of lactose induces expression of the genes required for lactose utilization n n Induction – stimulation of protein synthesis Inducer – molecule that stimulates synthesis Lactose – inducer of genes for lactose utilization 1950 s and 1960 s – Golden era of bacterial genetics n Advantages of E. coli and lactose utilization system n n n Culture large numbers of bacteria allow isolation of rare mutants Lactose genes not essential for survival (can use glucose as carbon source) Induction increases expression 1000 fold making mutant identification easy n Color changes using b-galactosidase enzyme (e. g. , OPNG, X-gal) make measurement of expression levels efficient
Coordinate repression and induction of three genes revealed by studies of lactose-utilization mutants n Jacques Monod and Francois Jacob – Pasteur Institute in Paris n Proposed Operon Theory of gene regulation n Single signal can simultaneously regulate expression of several genes that are clustered together on a chromosome and involve the same process n Because genes are clustered, they are transcribed together as single m. RNA n Clusters of genes are called Operons
Complementation Analysis of mutants identifies lactose utilization genes n n Monod et al. isolated many Lac- mutants unable to utilize lactose Complementation analysis identified three genes (lac. Z, lac. Y, and lac. A) in a tightly linked cluster Fig. 16. 5
Experimental evidence for repressor protein n Isolated mutant in lac. I gene Constitutive mutant – synthesized bgalactosidase and lac permease even in absence of lactose (inducer) n lac. I must be a repressor – cells must need lac. I protein product to prevent expression of lac. Y and lac. Z in absence of inducer n
Pa. Ja. Mo experiment n n n Fig. 16. 6 lac. I+, lac. Z+ x lac. I- lac. Z- FInitial synthesis of bgalactosidase stops Addition of inducer resumes synthesis Conclusion – initial lack of repressor allows synthesis. As lac. I is transferred, synthesis stops Repressor stops transcription by binding to operator site near promoter
Inducer releases repressor to trigger enzyme synthesis Addition of lactose inducer caused b – galactosidase synthesis to continue n Conclusion: Inducer binds to repressor so repressor can not bind to DNA n Allosteric effect - inducer bound to promotor changes conformation of protein so it can not bind to DNA n
Repressor has binding domains for operator and for the inducer Fig. 16. 7
Changes in the operator can also affect repressor activity Fig. 16. 8
Proteins act in trans DNA sites act only in cis Trans acting elements can diffuse through cytoplasm and act at target DNA sites on any DNA molecule in cell n Cis acting elements can only influence expression of adjacent genes on same DNA molecule n
Three experiments elucidate cis and trans acting elements using F’ plasmid n n Insert Figure 16. 9 a here Fig. 16. 9 a Inducible synthesis lac. I+ gene encodes a diffusible element that acts in trans by binding to any operator it encounters regardless of chromosomal location
Noninducible n Insert Figure 16. 9 b here n n Fig. 16. 9 b All operator sites (O+) eventually occupied by superrepressor lac. I supperrepressor can not bind inducer lac. Is mutant encodes a diffusible element that binds to operator regardless of chromosomal location (trans acting element)
Constitutive n Insert Figure 16. 9 c here n Fig. 16. 9 c Presence of O+ plasmid does not compensate for Oc mutation on bacterial chromosome Operator is cis acting element
The Operon Theory n The players n lacz, lac. Y, lac. Z genes that split lactose into glucose and galactose n Promotor site to which RNA polymerase binds n cis acting operator site n trans-acting repressor that can bind to operator (encoded by lac. I gene) n Inducer that prevents repressor from binding to operator Fig. 16. 10 a
The Operon Theory n Repression n n In absence of lactose, repressor binds to operator which prevents transcription Negative regulatory element Fig. 16. 10 b
The Operon Theory n Induction n Lactose present n n n Allolactose binds to repressor. Repressor changes shape and can not bind to operator RNA polymerase binds to promotor and initiates transcription of polycistronic m. RNA Fig. 16. 10 c
Positive control increases transcription of lac. Z, lac. Y, and lac. A n n n c. AMP binds to CRP (c. AMP receptor protein) when glucose is low CRP binds to regulatory region Enhances activity of RNA polymerase at lac promotor Fig. 16. 11
Some positive regulators increase transcription of genes in only one pathway n n Fig. 16. 12 Ara. C is a positive regulator for all arabinose genes which break down sugar arabinose Loss of function mutation results in little or no expression of genes
Molecular studies help fill in details of control mechanisms n Radioactive tag attached to lac repressor n n Repressor from lac. I+ cells purified and mixed with operator DNA, cosediment occurred Repressor from lac. I+ mixed with mutant operator DNA, no cosediment occurred Fig. 16. 13
Many DNA-Binding proteins contain a helix-turn-helix motif n n n Fig. 16. 14 a Two α-helical regions separated by a turn in the protein structure Helix-turn-helix motif fits into major groove of DNA Most repressor proteins
Specific amino acids in the a-helix determine the binding specificity of repressor proteins n Hybrid 434 -P 22 repressor engineered to have amino acid sequence that will bind to bacterial virus 434 and bacteriophage P 22 Fig. 16. 14 b
Most regulatory proteins are oligomeric § More than one binding domain §DNase footprint identifies binding region §DNase cannot digest protein covered sites Fig. 16. 15 a
The looping of DNA is a common feature of regulatory proteins n Ara. C acts as both a repressor and activator n No arabinose n n Arabinose present n Fig. 16 Binding to ara. O and ara. I 1 causes looping and prevents RNA from transcribing Ara. C binds to ara. I 1 and ara. I 2 bot not to ara. O. RNA polymerase interacts with ara. C at the ara. I sites and transcribes genes
How regulatory proteins interact with RNA polymerase n Negative regulators (lac repressor) n Physically block DNA-binding sites of RNA polymerase Positive regulators n Establish physical contact with RNA polymerase enhancing enzyme’s ability to initiate transcription n
Using the lac. Z gene as a reporter of gene expression Reporter gene – protein encoding gene whose expression in the cell is quantifiable by techniques of protein detection. n Fusion of reporter gene to cis acting regulatory regions allows assessment gene activity by monitoring amount of reporter gene product n
Fusion used to perform genetic studies of the regulatory region of gene X Fig. 16. 18 a
Creating a collection of lac. Z insertions in the chromosome Fig. 16. 18 b
Use of a fusion to overproduce a gene product Fig. 16. 18 c
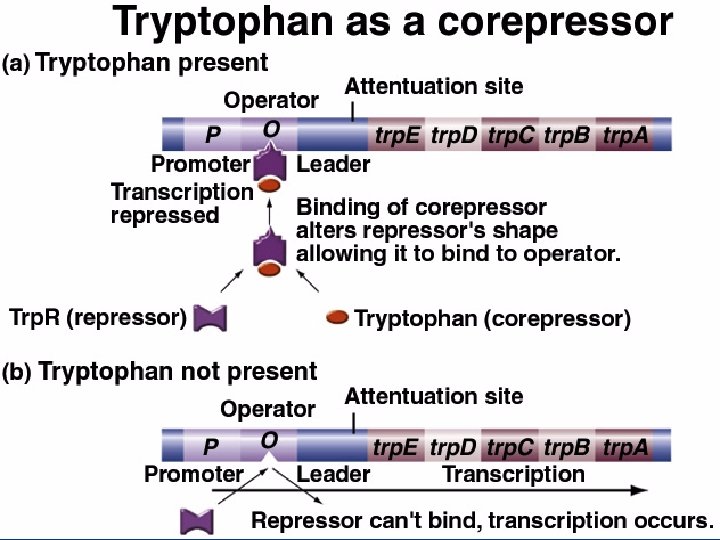
The attenuation of gene expression: Fine tuning of the trp operon through termination of transcription n The presence of tryptophan activates a repressor of the trp operon n trp. R gene produces repressor n Corepressor – tryptophan binds to trp repressor allowing it to bind to operator DNA and inhibit transcription
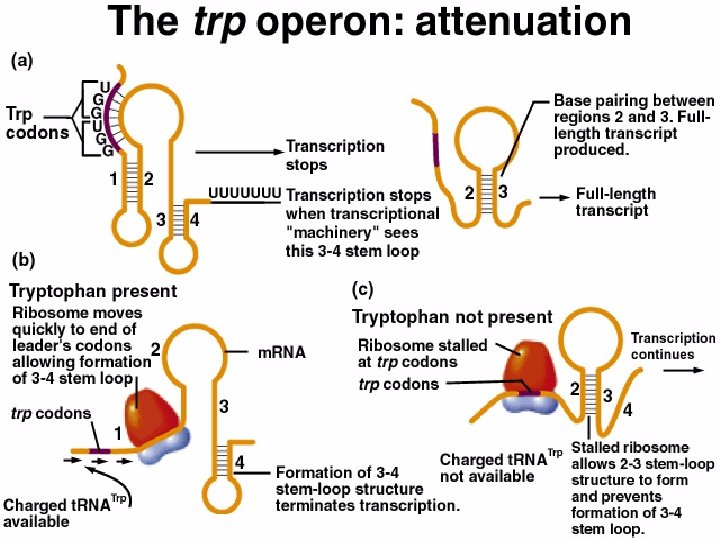
Termination of transcription fine tunes regulation of trp operon n trp. R- mutants are not constitutive n n Repressor independent change in trp expression Two alternative transcripts lead to different transcriptional outcomes n Leader sequence can fold in two different stable conformations n n Tryptophan present – ribosome moves quickly past codons in leader allowing stem-loop to form terminating transcription Tryptophan absent – ribosome stalls allowing normal stem loop structure to form and transcription proceeds normally
Global regulatory mechanisms coordinate the expression of many genes n n n Normal sigma factor (s 70) binds to RNA polymerase and recognizes sequence in promoter to initiate transcription Heat shock disables s 70 Product of rpo. H gene, s 32 binds to sequence in promoter of heat shock genes when heat stressed and starts transcription Fig. 16. 21 a
Factors influencing increase in s 32 activity after heat shock Increase in transcription of the rpo. H gene n Increase in the translation of s 32 m. RNA n n n stemming from greater stability of rpo. H m. RNA Increase in the stability and activity of the s 32 protein. Chaperones Dna. J/K bind and inhibit s 32 under normal conditions. At high temperature, binding to s 32 does not occur and more s 32 is free to associate with RNA polymerase. Inactivity of s 70 decreases competition with s 32 to form RNA polymerase holoenzyme
What enables transcription of s 32 during heat shock? n n Normal temperatures, rpo. H gene (encodes s 32) has promoter sequence recognized by s 70 which starts transcription High temperatures (no s 70) a different promoter sequence of the rpo. H gene is recognized by a different sigma factor, s 24
Summary n n n E. coli’s heat shock response is controlled by alternative sigma factors that recognize different promoter sequences Alternative sigma factors bind to RNA polymerase as temperatures change to start transcription of heat shock proteins The induction of alternative sigma factors that recognize different promoter sequences serve as global control regulatory mechanisms in E. coli and many other bacteria
Microarrays – a tool for uncovering changes in gene expression Cellular responses to global environmental changes can be measured by microarray analysis of m. RNA isolated from cultures grown in different environmental conditions n Comparisons of wild-type cultures with strains containing mutations in key regulatory regions help identify genes and regulatory elements involved in response to specific environmental changes n
Fig. 1. 13
Regulation of Virulence Genes in V. cholerae Bacterial agents of cholera sense changes in environment and transmit signals to regulators that initiate, enhance, diminish, or repress expression of various genes. n Three regulatory proteins – Tox. R, Tox. S, and Tox. T – turn on the genes for virulence n
Experiments generate model for regulation of virulence genes in V. cholerae Cloned two genes encoding subunits of cholera toxin: ctx. A and ctx. B n Made ctx. A-lac. Z reporter gene fusion n Created vector library of V. cholerae genomic DNA n Used E. coli to perform genetic manipulations n
n Isolated a gene that regulates expression of ctx operon n n Transformed E. coli containing ctx-lac. Z construct with clones containing V. cholerae DNA Clones that contain a positive regulator should turn on ctx-lac. Z construct Identified Tox. R, a transmembrane protein Identified Tox. S, helps Tox. R form dimers which helps it bind to DNA What genes does Tox. R regulate? n n n Gene fusions created to constitutive promoter Fusion introduced into strains of V. cholerae with lac. Z randomly inserted around the genome Identified intermediate regulator gene Tox. T, a transcriptional activator that binds to promoters of many genes, including ctx Tox. R/S or Tox. T can activate the ctx genes that produce toxin Tox. T alone activates additional virulence genes which encode pili and other proteins Transcription of Tox. T is regulated by Tox. R/S
Fig. 16. 22
Unanswered Questions Remain Why is there a cascade (Tox. R and Tox. T) of regulatory factors? n What DNA sequence in the promoters does Tox. R recognize? n What is the signal that’s makes the cholera bacteria start to colonize the small intestine? n How does Tox. R regulatory protein find binding sites on the chromosome? n