Gel di agarosio DNA una molecola carica negativamente





























- Slides: 35
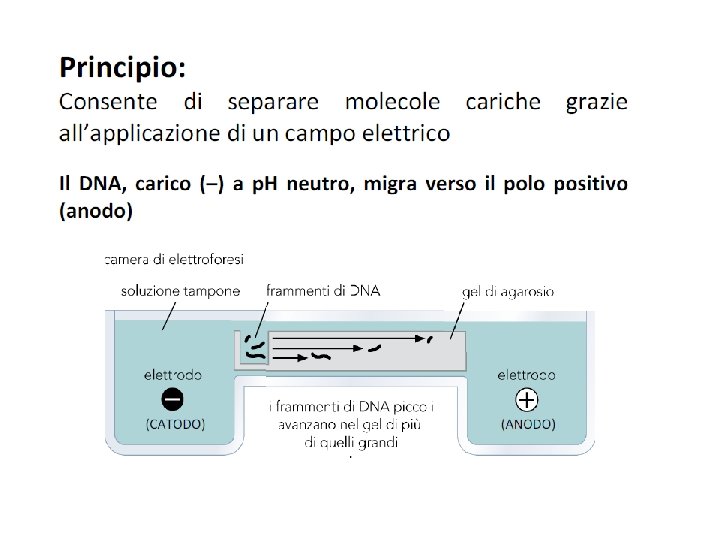
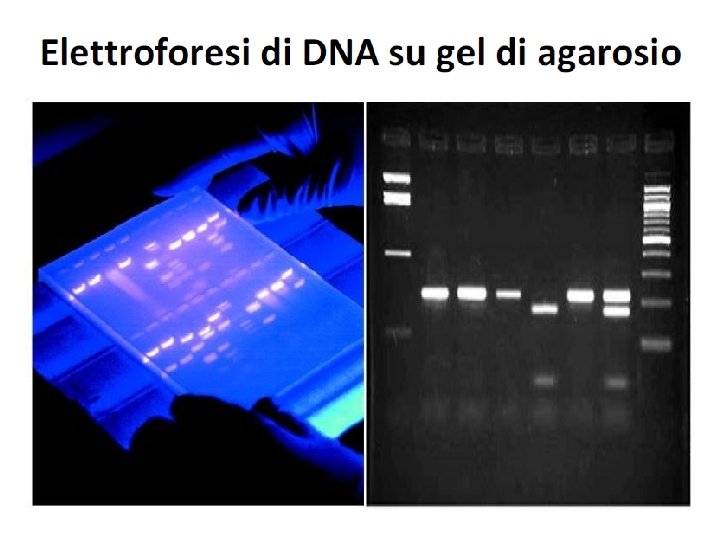
Gel di agarosio DNA è una molecola carica negativamente e quindi in un campo elettrico migra verso il polo positivo. La velocità di migrazione dipende da: 1. La sua forma 2. Il suo rapporto carica/massa Le molecole di DNA hanno la stessa forma e molto simili rapporti carica massa quindi si usa le gel elettroforesi che separa i frammenti di DNA in base alla loro dimensione
Elettroforesi di DNA Permette di separare, identificare e purificare frammenti di DNA. Contengono pori attraverso i quali il DNA migra al polo positivo. Più piccole sono le molecole, più veloce migrano sul gel Poliacrilammide DNA 5 -500 bp Alto potere risolutivo (1 bp) – Gel verticali – E costante Agarosio DNA 200 -50000 bp Varie concentrazioni. Minore potere risolutivo Gel orizzontali – E costante L’agarosio, in concentrazione opportuna, viene fuso insieme alla soluzione tampone. La soluzione fusa viene versata nella vaschetta e lasciata solidificare. Quando un campo elettrico viene applicato al gel il DNA, carico negativamente, si muove verso l’anodo.
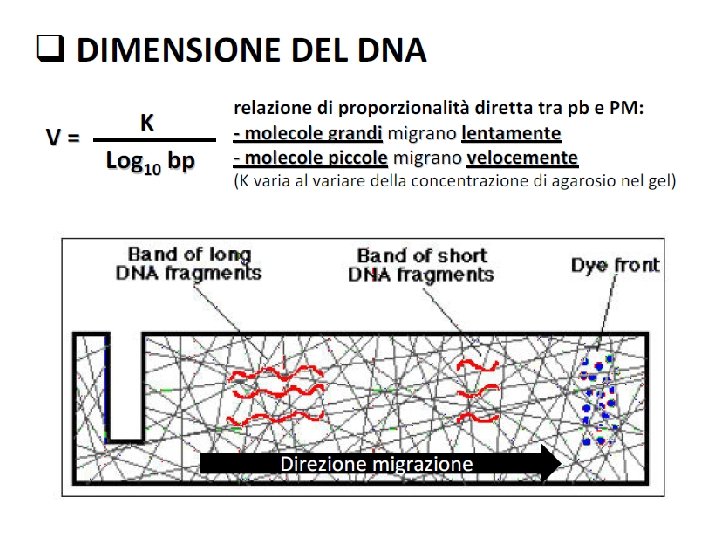
Fattori che influenzano la velocità di migrazione del DNA sul gel Dimensione della molecola di DNA La velocità di migrazione è inversamente proporzionale al lg 10 bp Concentrazione dell’agarosio Esiste una relazione lineare tra la concentrazione del gel ( ) e logaritmo della mobilità elettroforetica ( ) log = log °-Kr ° = mobilità elettroforetica libera Kr = coefficiente di ritardo (dipende dalle proprietà del gel e dalle dimensioni e forma delle molecole) Usando gel di conc. differenti è possibile separare una vasta gamma di molecole di DNA Conc. di agarosio Range di separazione (%) (Kb) 0. 3 5 -60 0. 9 0. 5 -7 1. 2 0. 4 -6 1. 5 0. 2 -3 2. 0 0. 1 -2
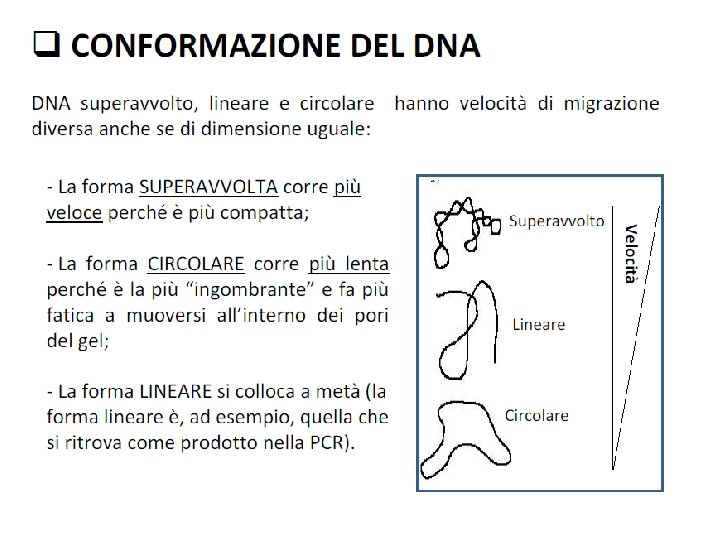
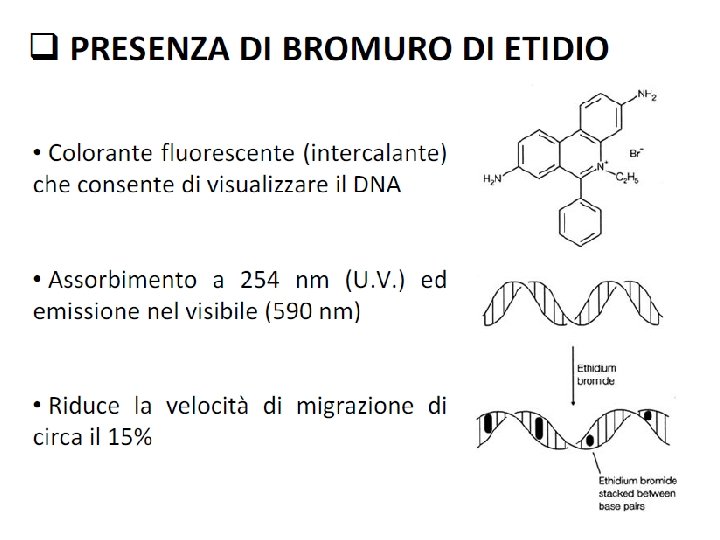
Fattori che influenzano la velocità di migrazione del DNA sul gel Conformazione del DNA Il DNA plasmidico puo’ trovarsi in forma superavvolta, parzialmente denaturata, o lineare. Le 3 forme, a parità di dimensioni, migrano con velocità diverse Voltaggio La velocità di migrazione è proporzionale al voltaggio applicato. Per avere ottima risoluzione il gel deve essere sottoposto ad un voltaggio 5 V/cm Composizione del tampone di corsa La presenza di ioni in soluzione permette la conducibilità elettrica e quindi la mobilità elettroforetica del DNA. Soluzioni tampone contengono EDTA (1 -2 m. M) p. H 8, Tris-borato(TBE)/Tris-acetato(TAE), 50 m. M. L’agarosio viene sciolto nello stesso tampone di corsa Loading buffer 6 x 0. 25 % blue di bromofenolo (migra alla stessa vel. di un frammento di 300 bp) 0. 25% xilene cianolo (migra alla stessa vel. di un frammento di 4000 bp) 30% glicerolo Bande corrispondenti ai frammenti di DNA si visualizzano utilizzando Et. Br (1 g/ml) un colorante fluorescente che si intercala alle basi del DNA e può essere rivelato attraverso luce UV. TBE 10 x p. H 8. 8 Acido borico Trizma base EDTA H 2 O fino a 0. 5 l 27. 5 g 162. 0 g 9. 5 g
Isolamento da gel del frammento della PCR Preparazione gel agarosio 2% Sciogliere 1. 0 g di agarosio in 50 ml TBE 1× scaldando su piastra. Quando l’agarosio è fuso (diventa limpido) aggiungere 5. 0 l di soluzione di Et. Br 10 mg/ml (già pronta, maneggiare con guanti). Versare la soluzione tiepida nella vaschetta da gel in cui è stato inserito il pettine. Caricare su gel i seguenti campioni: 1) 100. 0 l frammento PCR + 20. 0 l loading buffer 2) 6. 0 l marker (100 bp) containing loading buffer Far correre a 100 V per 30 -40’. Osservare la banda corrispondente al frammento di gene amplificato corrispondente a 473 bp. Tagliare con il bisturi il pezzetto di gel contenente la banda e trasferirlo in un eppendorf sterile (dopo aver determinato la tara di quest’ultimo). Estrarre il DNA dal gel con il QIAquick Gel extraction kit.
Preparazione di un gel di agarosio
Estrazione del DNA da gel Il prodotto viene estratto da gel con il QIAquick gel extraction kit. Si utilizza per DNA da 70 bp a 10 Kb. In ogni colonna si possono processare fino a 400 mg di agarosio. Si basa sullo stesso principio del protocollo di purificazione della PCR. In questo caso il gel deve essere fuso in precedenza in presenza di una soluzione (buffer QG) che solubilizza il gel e assicura il successo legame del DNA alla membrana di gel di silice.
Estrazione del DNA da gel
Digestione del prodotto della PCR con enzimi di restrizione Hind. III 5’-attcccaagctttagcgatggcgacgaaggc… 3’-taagggttcgaaatcgctaccgctgcttccg. . …………………. gatcgcccaataagtcgacgcgtgacc-3’ ctagcgggttattcagctgcgcactgg– 5’ Sal. I vettore di espressione Il prodotto della PCR deve essere tagliato con gli enzimi di restrizione prima di essere clonato nel vettore p. PHSOD 1 Iq, tra i siti Hind. III e Sal. I Hind. III gene Sal. I
Digestione del prodotto della PCR con enzimi di restrizione Digestione del prodotto della PCR con gli enzimi Hind. III e Sal. I H 2 O Buffer 2 (10 ) Frammento PCR Hind. III (20 U/ml) 14. 0 l 5. 0 l 30. 0 l 1. 0 l 50. 0 l O/N 37°C H 2 O Buffer x Sal (10 ) Frammento PCR tagliato BSA 100 10 mg/ml Sal. I (20 U/ml ) O/N 37°C 38. 5 l 10. 0 l 50. 0 l 0. 5 l 1. 0 l 100. 0 l
Tabella di compatiblità per la doppia digestione seq = sequenziale
Digestione enzimatica “close to the end of DNA fragments”
La ligasi Cosa fa: Quando si usa nel gene cloning:
La ligasi Scarsa efficienza perché la ligasi è incapace di tenere unite le sue molecole di DNA, quindi dobbiamo avere alta conc. di DNA per avere alta resa Molto più efficiente perché si forma un o stabile intermedio sul quale la ligasi può reagire. Più lunghe sono le sticky ends e maggiore è l’efficienza della reazione
La DNA ligasi più utilizzata è quella codificata dal batteriofago T 4, un enzima, ATPdipendente, ottenuto da cellule di E. coli infettate dal fago T 4. La T 4 DNA ligasi è capace di legare tra loro estremità coesive e, seppur a minor efficienza, è capace di ligare tra loro anche estremità piatte. La DNA ligasi di E. coli, un enzima NAD+-dipendente presente in E. coli, può legare tra loro soltanto estremità coesive ed è relativamente poco utilizzata. Le estremità “coesive”, che contengono cioè corti filamenti a singolo filamento complentari tra loro, che vengono generate da molti enzimi di restrizione, facilitano il compito della ligasi, perché le estremità tendono ad appaiarsi tra loro. In queste condizioni le ligazioni si effettuano generalmente tra 4 e 37°C, per un tempo variabile tra 1 e 24 ore. Se, invece, bisogna ligare, estremità “blunt” la ligazione è più difficile e si effettua a bassa temperatura e a più elevata concentrazione di enzima e di frammenti da legare per favorire l’incontro delle molecole da legare.
Per l’attività della ligasi sono necessari: • ATP • Un gruppo fosfato al 5’ • Un gruppo -OH al 3’ Il meccanismo della ligazione avviene in 3 fasi: 1) 2) 3) La ligasi reagisce con l’ATP per formare un complesso covalente ligasi-AMP La ligasi-AMP reagisce con il P al 5’ e con una reazione di transesterificazione lega l’AMP al fosfato rilasciando l’enzima Il gruppo -OH reagisce con il 5’P e e determina la formazione di un nuovo legame fosfodiesterico
Ottimizzazione della ligazione Per ottenere buoni risultati la ligazione deve essere accuratamente ottimizzata rispetto alla: • temperatura e al tempo di reazione • alla concentrazione totale del DNA • alla concentrazione dell’inserto e del vettore. Riguardo alla temperatura bisogna considerare che si contrappongono due aspetti opposti: la stabilizzazione dell’appaiamento tra estremità coesive, ottimale a basse temperature, e l’attività enzimatica della ligasi, massima a 37°C. Come compromesso si utilizza spesso una temp. di 16°C per 12 ore. Per quanto riguarda la concentrazione è importante sia quella del DNA totale T , che quella dell’inserto I e del vettore V. In linea di massima basse concentrazioni di DNA favoriscono le reazioni di primo ordine come la ricircolarizzazione del vettore, la cui velocità è linearmente proporzionale alla sua concentrazione. D’altra parte, aumentare la concentrazione dell’inserto aumenta la probabilità di avere vettori con inserti multipli.
Fosfatasi alcalina Per ottimizzare la reazione di ligazione, nel senso di evitare una elevata frequenza di ricircolarizzazione del vettore. P-5’ AATTC HO-3’ vettore inserto P-5’ AATTC HO-3’ 3’OH TTAAG -5’P vettore 3’OH TTAAG -5’P Una strategia per superare questo problema consiste nell’utilizzo di una fosfatasi, come ad esempio la fosfatasi alcalina (BAP), un enzima che rimuove il gruppo fosfato al 5’impedendo così l’azione della ligasi. La defosforilazione del vettore con una fosfatasi impedisce la ricircolarizzazione del vettore, abbassando sensibilmente il background. Poiché il vettore defosforilato non può essere ligato, inoltre, è possibile utilizzare un eccesso molare di vettore, minimizzando la possibilità di avere inserzioni multiple.
P-5’ AATTC HO-3’ vettore 3’OH fosfatasi TTAAG -5’P HO- AATTC HO-3’ vettore 3’OH TTAAG-OH Sebbene un vettore defosforilato non possa essere circolarizzato dalla ligasi, può essere ligato ad un inserto sfruttandone il suo fosfato al 5’. L’assenza di fosfati al 5’ del vettore produrrà due nicks, uno per filamento. Queste interruzioni a singolo filamento, tuttavia non influenzeranno negativamente la stabilità del costrutto (vettore + inserto) e saranno riparate dai sistemi di riparo di E. coli subito dopo la trasformazione. P-5’ AATTC HO-3’ vettore inserto HO- AATTC HO-3’ 3’OH TTAAG -5’P vettore 3’OH TTAAG-OH + ligasi vettore OH HO inserto OH HO vettore
Quando frammenti di DNA sono blunt-ended, Come si procede? Approccio che utilizza linkers:
Quando frammenti di DNA sono blunt-ended, Come si procede? Approccio che utilizza adattatori:
Quando frammenti di DNA sono blunt-ended, Come si procede? La Ligasi è incapace di formare legami fosfodiesterei tra 5’-OH e 3’-OH
Produzione di sticky ends attraverso una coda omopolimerica Addizione di deossiguanosina attraverso un enzima deossinucleatidil transferasi che aggiunge nucleotidi sul 3’-OH terminale di una molecola di DNA double stranded. La reazione in (c) non è necessaria nel test tube se l’omopolimero è lungo più di 20 nucleotidi perché è stabile abbastanza da essere introdotto nella cellula batterica ospite dove viene riparata da DNA polimerasi e ligasi
Plasmide di espressione della SOD Ampicillina Sequenza Nterminale per far esprimere la proteina periplasmatica
Preparazione del vettore di espressione Digestione vettore H 2 O sterile Buffer 2 (10 ) Vettore p. PHSOD 1 Iq (500 ng/ l) Hind. III (20 U/ l) >1 h, 37 °C H 2 O sterile Buffer x Sal (10 ) Bsa (100 g/ l) Vettore tagliato con Hind. III Sal. I(20 U/ l) >1 h, 37 °C 12. 0 2. 5 10. 0 0. 5 25. 0 l l l Defosforilazione vettore 19. 0 l 5. 0 l 0. 5 l 25. 0 l 0. 5 l 50. 0 l totale H 2 O sterile Buffer 10 AP Vettore tagliato AP (1 U/ l) 39. 0 l 10. 0 l 50. 0 l 1. 0 l 100. 0 l 30’, 37 °C Per disattivare l’enzima: 15’, 65 °C AP = Fosfatasi alcalina che rimuove fosfato al gruppo 5’ del DNA
Isolamento da gel del vettore Preparazione gel agarosio 1% Sciogliere 0. 5 g di agarosio in 50 ml TBE 1× scaldando su piastra. Quando l’agarosio è fuso (diventa limpido) aggiungere 5. 0 l di soluzione di Et. Br 10 mg/ml (già pronta, maneggiare con guanti). Versare la soluzione tiepida nella vaschetta da gel in cui è stato inserito il pettine con i denti uniti a due. Caricare su gel i seguenti campioni: 1) 6. 0 l marker (1000 bp) containing loading buffer 2) 100. 0 l vettore defosforilato + 20. 0 l loading buffer 3) 1. 0 l vettore p. PB 10 non tagliato + 4. 0 l H 2 O + 1. 0 l loading buffer Far correre a 100 V per 30 -40’. Osservare la banda corrispondente al frammento di vettore di 5109 paia di basi. Tagliare con il bisturi il pezzetto di gel contenente la banda e trasferirlo in un eppendorf sterile (dopo aver determinato la tara di quest’ultimo). Pesare l’eppenderf con il DNA
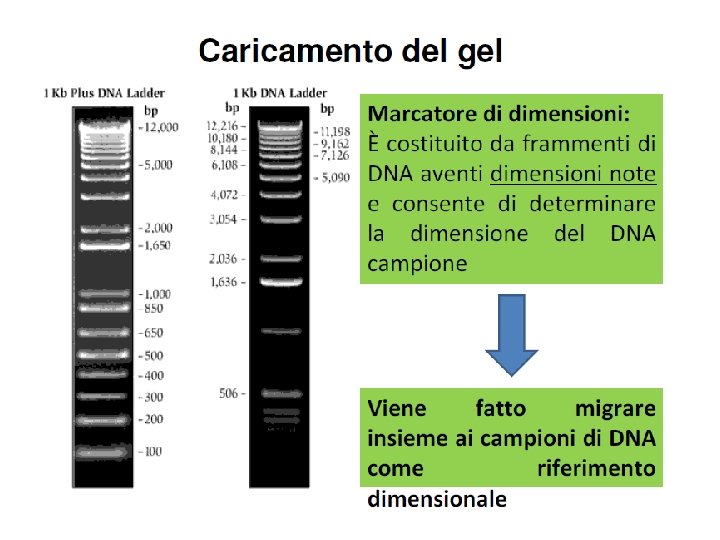
Determinazione della concentrazione del DNA La concentrazione di una campione di DNA puo’ essere fatta in 2 modi 1) Valutazione dell’intensita’ della banda su gel di agarosio e confronto con standard Quando DNA conc. è minore di circa 20 ng intensità della banda con Et. Br è debole o non visibile 2) Per via spettrofotometrica. 50 g/ml DNA a doppio filamento Abs 260 = 1 40 g/ml DNA a singolo filamento Abs 260 = 1 Il rapporto tra Abs 260/Abs 280 fornisce una stima della purezza Campione puro Abs 260/Abs 280 1. 8 Purine assorbono < 260 nm; pirimidine = 260 nm
Quantificazione su gel del frammento e del vettore Preparazione gel agarosio 2% Sciogliere 1. 0 g di agarosio in 50 ml TBE 1× scaldando su piastra. Quando l’agarosio è fuso (diventa limpido) aggiungere 5. 0 l di soluzione di Et. Br 10 mg/ml (già pronta, maneggiare con guanti). Versare la soluzione tiepida nella vaschetta da gel in cui è stato inserito il pettine. Caricare su gel i seguenti campioni: 1) 4. 0 l frammento PCR + 6. 0 l H 2 O + 2. 0 l loading buffer 2) 4. 0 l vettore + 6. 0 l H 2 O + 2. 0 l loading buffer 3) 1. 0 l marker (100 bp) + 4. 0 l H 2 O + 1. 0 l loading buffer 4) 1. 0 l marker (1 Kbp) + 4. 0 l H 2 O + 1. 0 l loading buffer Confrontando l’intensità delle bande 1) e 2) con quelle presenti nel campione 3), a quantità nota, valutare la concentrazione del frammento PCR e del vettore.
Reazioni di ligasi tra frammento e vettore Si usano 20. 0 ng di vettore. La reazione, catalizzata dall’enzima T 4 DNA ligasi avviene a p. H 7. 5, in presenza di Mg. Cl 2 e ATP La quantità di frammento da clonare viene stimata tenendo conto che si useranno frammento e vettore nei rapporti molari 5: 1 e 20: 1. Vettore 5109 bp Frammento 473 bp 20 ng : X = 5109 : 473 X (ng di frammento) = (20 x 473)/5109 = 1. 85 ng 5 X = 9. 2 ng 20 X = 37. 0 ng >