Fotosinteza Zshema linearni transport elektrona Linearni transport elektrona


 Ciklični transport elektrona samo ATP Pseudociklični transport elektrona samo")


 Askorbat-glutationski ciklus (Halliwel-Asada ciklus) Regeneracija Asc i")
 Regeneracija Asc i potrošnja NADPH")



 Prvi produkt nije saharoza")
 Karboksilacija – vezivanje CO 2 u organsku materiju, za ribulozo-1,")







 u saradnji tri organele 2 glikolata (4 C) u peroxizom")




,")
 •")






 u saradnji tri organele GS-GOGAT")


 Prethodno vezivanje CO")









- Slides: 47
Fotosinteza
Z-shema linearni transport elektrona
Linearni transport elektrona NADPH+ATP (2+3) Ciklični transport elektrona samo ATP Pseudociklični transport elektrona samo ATP O 2 • O 2 Fd
http: //vcell. ndsu. edu/animations/photosynthesis/movie-flash. htm http: //www. dailymotion. com/video/x 2 fkksu
Višak energije i mehanizmi zaštite od oštećenja Fe 3+ + • O 2− → Fe 2+ + O 2 Fe 2+ + H 2 O 2 → Fe 3+ + OH− + • OH Fenton • O 2 - + H 2 O 2 → • OH + OH- + O 2 Haber-Weiss
Ksantofilni ciklus “gašenje” pobuđenog stanja hlorofila (quenching) Askorbat-glutationski ciklus (Halliwel-Asada ciklus) Regeneracija Asc i potrošnja NADPH Ciklus voda-voda Uklanjanje ROS
Ciklus voda-voda Uklanjanje ROS Askorbat-glutationski ciklus (Halliwel-Asada ciklus) Regeneracija Asc i potrošnja NADPH
Fotosinteza je ograničena koncentracijom CO 2 i intenzitetom svetlosti. Temperatura utiče samo na deo fotosinteze koji zavisi od CO 2 dok nema uticaj na deo koji zavisi od svetlosti
faze fotositeze svetla i tamna fotohemijska i biohemijska • Fotohemijska faza podrazumeva • Apsorpciju svetlosti i oksidaciju hlorofila u RC • Redukciju H 2 O i oslobađanje O 2 • Kretanje elektrona kroz redoks lanac • Redukciju NADP+ • Sintezu ATPa
Biohemijska faza fotosinteze Koriste se ATP i NADPH sintetisani tokom fotohemijske faze
Berkley, Amerika 40 -ih i uvođenje radioaktivnih izotopa (14 C) Prvi produkt nije saharoza Prvi stabilan intermedijer je 3 -fosfoglicerat Brzo se javlja još veliki broj jedinjenja
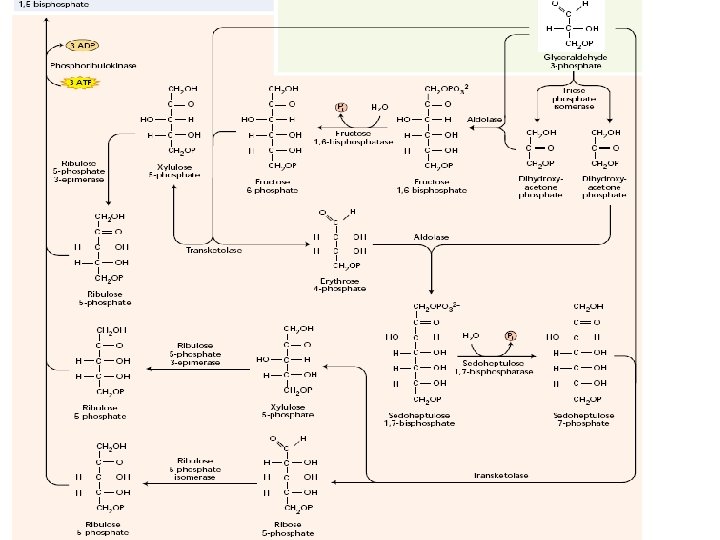
Kalvinov ciklus (reduktivni PPC) Karboksilacija – vezivanje CO 2 u organsku materiju, za ribulozo-1, 5 -bisfosfat Redukcija – od oksidovanog (C+4) do redukovanog (C+1) u GAP Regeneracija akceptora – Ru-1, 5 -b. P
Rubis. CO • Jedini enzim koji omogućuje fiksaciju atmosferskog CO 2 za izgradnju biomase • Do 50% solubilnih proteina lista • Najzastupljeniji protein na Zemlji • Afinitet prema CO 2 je dovoljno visok da enzim radi i pri niskim koncentracijama CO 2
• Fosforilacija fosfoglicerata • Redukcija bisfosfoglicerata • NADPH GA-3 -P dehidrogenaza • Enzim sličan citosolnom koji učestvuje u glikolizi
Da bi tekao ciklus neophodna je regeneracija akceptora – Ru. BP Faza indukcije Samo 1/6 triozo fosfata se koristi za sintezu skroba i saharoze Brzina ciklusa zavisi od koncentracije intermedijera (lag faza pri uključivanju osvetljenja)
Foton ATP NADPH Zemljište Atmosfera Voda Neposredan izvor energije Osnovni izvor energije Energija Redukcioni potencijal Materija
1 CO 2 – 8 fotona – 8*172 k. J 6 mol CO 2 - 1 mol fruktoze - 8400 k. J Oksidacija fruktoze – 2800 k. J Za svaki CO 2 je potrebno 2 NADPH i 3 ATP Za jedan molekul fruktoze treba 12 NADPH i 18 ATP To je 12*217 k. J (oksidacija NADPH) + 18* 29 k. J (hidroliza ATP) 3126 k. J –ef ikasnost 90%
Rubis. CO • Jedini enzim koji omogućuje fiksaciju atmosferskog CO 2 za izgradnju biomase • Do 50% solubilnih proteina lista • Najzastupljeniji protein na Zemlji • Afinitet prema CO 2 je dovoljno visok da enzim radi i pri niskim koncentracijama CO 2 • Katalizuje i oksigenaciju • Afinitet za O 2 manji nego za CO 2
Ribulozo bisphosphate Carboxylase Oxigenase • CO 2: O 2 u atmosferi je 0. 039: 21 • Km [CO 2] (9µM)<Km[O 2] (535µM) • Kcat [CO 2] (3. 3/s)>Kcat[O 2](2. 4/s) • Sa povišavanjem temperature menja se CO 2/O 2 specifičnost kao i rastvorljivost gasova
Fotorespiracija (C 2 put) u saradnji tri organele 2 glikolata (4 C) u peroxizom GS-GOGAT Transformacija u glicin, koji se exportuje u mitohondriju gde se od dva Gly sintetiše Ser (3 C) Ser u peroxizom gde se transformiše u glicerat Glicerat u hloroplast, fosforiliše se i uključuje u Kalvinov ciklus HPR CAT transaminaza Glutamin sintetaza, Glutamat sintaza ( NADPH-dependent glutamine: 2 oxoglutarate aminotransferase) 3 C se vraćaju u hloroplast CO 2 se oslobađa u mitohondrijama i može ponovo da se asimiluje Gly dekarboksilaza kompleks N se reasimiluje i značajno doprinosi sintezi ak Ser hidroximetil transferaza
2 Ru. BP + 3 O 2 + 3 H 2 O 3 3 PGA + CO 2 + Pi Gubitak za fotosintezu (1/4 C) – smanjuje produktivnost biljaka Posledica karakteristika Rubis. CO, odnosa CO 2/O 2 i temperature Fotorespiracija NIJE ISTO što i respiracija Zašto postoji? • Mehanizam katalize • Zaštita fotosintetičkog aparata – rasipanje viška ATP i NADPH
Regulacija Kalvinovog ciklusa Visoka efikasnost znači da postoji precizna regulacija Više nivoa i mehanizama regulacije koji zajedno obezbeđuju efikasnost procesa Genska ekspresija (jedarna i hloroplastna) – količina enzima Posttranslacione modifikacije – kinetički parametri enzima kovalentne modifikacije (S-S; karbamilacija) modifikacije nekovalentnih veza (p. H, jonska jačina; asociranje sa tilakoidima) Glavni faktor koji reguliše ove procese je SVETLOST • Rubis. CO • NADP: GA 3 P dehidrogenaza • Fruktozo-1, 6 -bisfosfataza • Sedoheptulozo-1, 7 -bisfosfataza • Ribulozo-5 -fosfat kinaza • ATP sintaza
Svetlosna regulacija aktivnosti enzima • NADP: GA 3 P dehidrogenaza • Fruktozo-1, 6 -bisfosfataza • Sedoheptulozo-1, 7 -bisfosfataza • Ribulozo-5 -fosfat kinaza • ATP sintaza (γ subjedinica) na aktivnost enzima utiču i p. H [Mg++] odnos NADPH/NADP (kompetitivna inhibicija)
Regulacija aktivnosti Ru. Bis. CO 8 malih subjedinica kodiranih u jedru 8 velikih subjedinica kodiranih u plastidu Aktivni centar je na dodiru dve velike subjedinice Aktivacija CO 2 (karbamilacija ε-NH 2 Lys u aktivnom centru) a zatim karbamat vezuje Mg++ Zatim se vezuje drugi molekul CO 2 u aktivnom centru koji biva inkorporiran u organsku materiju
Ova aktivacija je sprečena ako je u aktivnom centru vezan fosfatni šećer (Ru. BP), koji se uklanja Rubis. CO aktivazom, koju kod nekih vrsta aktivira feredoksin-tioredoksin sistem Analog 2 -karboksi 3 -keto arabinitol 1, 5 bisfosfata
Enzimske aktivnosti reguliše i svetlosno-zavisan fluks jona (promene p. H i oslobađanje Mg++) • Rubis. CO • Fruktozo-1, 6 -bisfosfataza • Sedoheptulozo-1, 7 -bisfosfataza • Ribulozo-5 -fosfat kinaza
Regulacija reduktivnog PPC i oksidativnog PPP
Oksidativni PPP Redukcioni potencijal za reakcije u mraku Održava nivo prekursora Ru. BP Ribozo-5 -fosfat (sinteza nukleinskih kiselina) Eritrozo-4 -fosfat (+PEP) (sinteza fenolnih jedinjenja) Sinteza intermedijera Kalvinovog ciklusa u mladim listovima
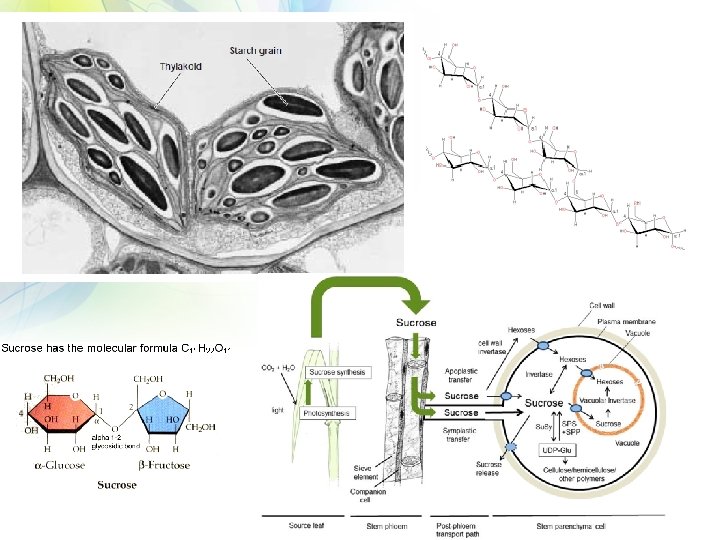
Primarni produkti fotosinteze su Sekundarni produkti fotosinteze
Skrob – glavni rezervni oblik
Saharoza – glavni transportni oblik
3 PGA+ Pi- TRX F 26 b. P, AMP G 6 P+ Pi- Sinteza saharoze ili glikoliz
Fotorespiracija (C 2 put) u saradnji tri organele GS-GOGAT
Mehanizmi koncentrovanja CO 2 U današnjoj atmosferi, kao i u vodenoj sredini, nije dovoljna koncentracija CO 2 da bi Rubis. CO radio maksimalnom brzinom Alge i bakterije imaju CO 2 i HCO 3 - pumpe
Fotorespiracija zavisi od temperature
C 4 i CAM metabolizam Normalan C 3 metabolizam (Calvinov ciklus) Prethodno vezivanje CO 2
C 4 i CAM metabolizam C 3 C 4 CAM stome Danju otvorene, noću zatvorene δotvorene danju, δ zatvorene noću Danju zatvorene, noću otvorene CO 2 Fiksacija u C 3 jedinjenje Fiksacija u C 4 organsku kiselinu voda Podložne prekomernom gubitku Manje podložne gubitku vode Ne gube vodu, mala produktivnost C 4 – prostorno razdvajanje (prethodne) fiksacije CO 2 i Kalvinovog ciklusa CAM – vremensko razdvajanje (prethodne) fiksacije CO 2 i Kalvinovog ciklusa
C 4 biljke Ćelije bez hloroplasta Krupni agranalni hloroplasti Biljke tropskih predela (otkrivena kod tropskih trava) 16 familija monokotila i dikotila Polifiletsko poreklo Paralelna evolucija strukture i funkcije
Prvi produkt nije 3 PGA već malat, oksalacetat ili aspartat Mezofilne ćelije sadrže PEP karboksilazu i piruvat-ortofosfat dikinazu Kranc ćelije sadrže enzime Kalvinovog ciklusa
1 2 4 3 1. 2. 3. 4. Karboksilacija PEP i sinteza malata i/ili aspartata u mezofilnim ćelijama Transport C 4 kiseline u ćelije sare Dekarboksilacija C 4 i oslobađanje CO 2 u ćelijama sare Transport C 3 (piruvat ili alanin) u mezofilnu ćeliju za regeneraciju PEP
3 tipa C 4 metabolizma: 1. NADP-malatni 2. NAD 3. PCK-aspartatni
Svetlosna regulacija TRX fosforilacija ADP-fosforilacija
Potrebna su dva dodatna ATPa za prethodnu fiksaciju, tako da je ukupan utrošak Kalvinovog ciklusa 5 ATP i 2 NADPH Ovo je doska “skupo” (skoro kao fotorespiracija), ali omogućuje efikasnu fotosintezu u nepovoljnim uslovima, pa je u nepovoljnim uslovima produktivnost veća nego kod C 3 biljaka Benefit C 4 metabolizma obezbeđuje visok afinitet PEP karboksilaze za CO 2, i nemogućnost oksigenacije To omogućava i da stome budu delimično zatvorene, pa biljke gube manje vode
CAM biljke Sukulente iz različitih familija Konzervacija vode – 100 - g/g. CO 2 (vs 500 g/g. CO 2 kod C 3 I 300 kod C 4) Noću otvorene stome Krupna vakuola
Noćno zakišeljavanje lista Danju povišenje CO 2 u intercelularima PEP karboksilazu inhibira malat, aktivira G 6 P Kod CAM aktivna noću (diurnalni ritam), kod C 4 aktivna danju (svetlost)