Fossils and systematics Intended learning outcomes n n





 n n In a devastating critique, Colin Patterson (1981) tried to assess")
















 chinensis")
![“Probably no-one will now deny that trilobites are crustaceans” Raymond (1920) “The Xyphosurida [Chelicerata]](https://slidetodoc.com/presentation_image_h/d616590587f1af9c68ffef6c4ad72a1c/image-27.jpg "“Probably no-one will now deny that trilobites are crustaceans” Raymond (1920) “The Xyphosurida [Chelicerata]")
 sudden appearance of fully-formed taxa, lots")
 sudden appearance of fully-formed taxa, lots")
")








 • We expect old groups to be morphologically distinct, because they have")










 • All fossils lie in stem-groups. • Old fossils lie in deep")
- Slides: 50
Fossils and systematics
Intended learning outcomes n n Describe the basis for cladistics in terms of congruence and parsimony, and describe the four types of parsimony. Construct simple character matrices and assess short trees for length and other simple statistics. Apply the stem and crown group concepts to fossil taxa, and give examples of their use in the Cambrian explosion, with named taxa. Describe the ways in which fossils can affect systematics, with examples.
Old views n Fossils could be used to simply ’see’ ancestry. E. g. the views of G. G. Simpson etc in the 1940’s and 1950’s. (But Darwin already foresaw some of the problems).
Problems with old view n n n A whole set of characteristic problems started to appear. Despite extensive documentation of many fossils, evolutionary history for many groups seemed still highly problematic. Typically, the problem was typified by the appearance of very large numbers of high-level systematic groups (Orders, Classes etc) with no connection between them, often for tiny numbers of taxa (e. g. one-species Classes and so on).
Patterson (1981) n n In a devastating critique, Colin Patterson (1981) tried to assess (from a very critical standpoint) the influence fossils had in fact had on systematics. His answer: virtually none, despite looking at a whole range of different taxa – fish, arthropods etc. Patterson (1977) and Nelson (1978) argued that fossils were totally reliant on living groups for their interpretation, and thus could not be used in a primary sense for systematics: rather, they could only be ’slotted in’ after a systematic analysis. Fossils were argued to be imperfectly preserved with many missing characters that could only be filled by inference from living forms.
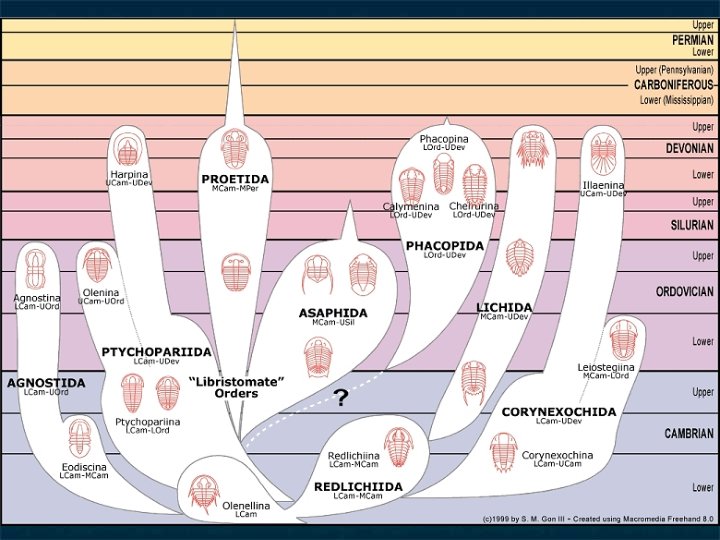
An example of pre-cladistic analysis: the arthropods of the Burgess Shale n n Previous analyses: e. g. Størmer; Whittington. Typical levels of high-level, lowdiversity taxa.
Given the problems of old-style systematising of fossils, are they any use at all in systematics? n n The cladistic onslaught led to a reappraisal of systematics in fossils. Question now is: given the correct approach, can fossils provide important information? Empirical tests involving leaving in/deleting fossils ’total evidence’ analysis Simulation experiments testing parsimony etc
Are fossils really of no use in phylogeny? n Famous papers: Doyle and Donoghue (1987); Gauthier et al. (1988) tested the importance of fossil evidence in seed plants and tetrapods. n Technique: pretend fossils had never been discovered, and code living taxa accordingly (important for homology assessment…). Compile data matrix, and then delete some/all fossil taxa to compare successive results. n Results: inclusion of fossils decisively influenced tetrapod relationships; and in seed plants changed character polarities etc. n Later studies have tended to confirm this view, e. g. Novacek (1992) on mammals.
Gauthier et al.
Total Evidence n n More data from fossils must improve the reliability of the tree. Recent taxa can only provide a small sample of all the character combinations that have existed. ”Every feature of a fossil, no matter how strange, has in the end to be explained” (Dick Jefferies).
Missing Evidence? n n Some have argued that fossils have many missing characters, and thus add vastly to the number of most parsimonious trees; their addition in general to analyses will destabilise them: fossils should therefore be excluded. However, ’missing data’ is a problem for recent taxa too! Relative number of characters is not so important to analyses as the importance of the characters that are there (e. g. Cloutier on Sarcopterygian fish). It is therefore not justifiable to remove taxa on the grounds of incompleteness – the characters that are present may be particularly significant.
Simulation studies n n n Why might fossils be particularly important? Essentially, because they break up long branches and reduce the problem of longbranch attraction. May be particularly important for groups such as phyla that have a very long separate evolutionary history, i. e. groups with long stem-groups (see later…)
Effects of fossils on systematics n n Can have three effects Can reveal a character in the past had a broader distribution than it does now (i. e. can reveal that supposed apomorphies are in fact symplesiomorphies). Can demonstrate that supposed synapomorphies are in actuality homoplasies. E. g. phalanges in horses and hyracoids - basal fossils in hyracoids have only the plesiomorphic condition, strongly suggesting convergence between the two. Fossils can provide intermediate character states that show putatively different characters to be in actuality states of the same character. E. g. : the mammalian middle ear bones compared to the postdentary bones of the reptilian lower jaw.
Dating and stratigraphy n n Old assumption was that old fossils were ’more primitive’ – i. e. if two fossils had different character states of the same character, the older fossil possessed the primitive character state, and the younger fossil the more derived state. Nothing is more likely to annoy cladists than this assertion! E. g. ’The general consensus is that using the fossil record to establish polarity or to test theories of relationship is suspect, since any false or unwanted results may be explained away by invoking incompleteness of the record’ (Forey 1992)
Conclusions about stratigraphy n n 1. Cloutier and others have shown, as one might expect, that old fossils have more plesiomorphic character states than younger ones, and fewer uncodable ones. 2. Fossils with more plesiomorphic characters are more likely to break up long branches and thus tackle long branch attraction: they may also undermine character optimization based on extant taxa.
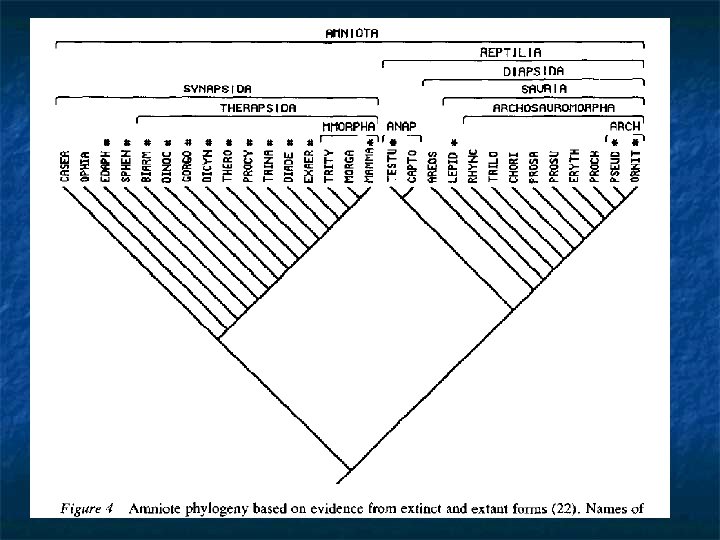
Systematisation of fossils n n n Consider the relationship of fossil taxa to living ones. Questions to ask: do all fossils fit into extant taxa? Consider a pair of sister taxa monophyletic living groups. First, look at where the apomorphies of each group accumulate: in the group, or outside them? Obviously outside them, or they would not be shared by all the members of the group! Second, look at the last common ancestor of the two groups. Again, is it a member of either group? Obviously not, as by hypothesis, the groups are both monophyletic. CONCLUSION: Some organisms exist that fall outside the scope of extant groups at a particular level.
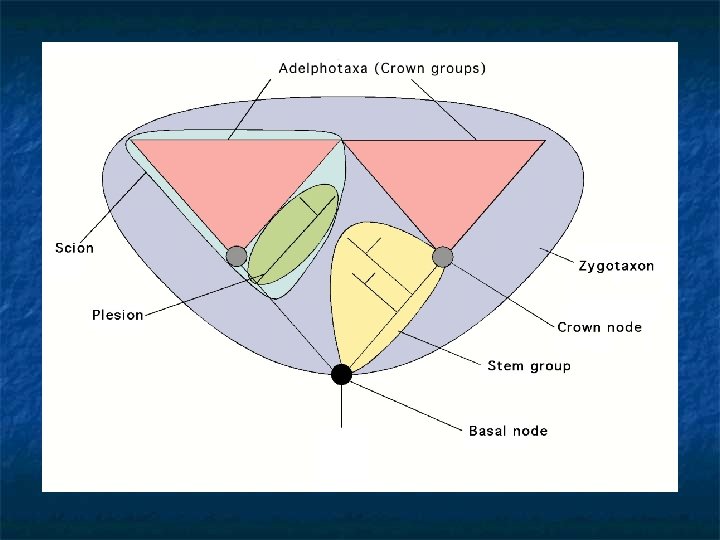
More on Stem and Crown groups n n n Oddly enough, ALL fossils fall into a stem-group at one level or other of the hierarchy. Stem-groups are always paraphyletic: but unlike most paraphyletic groups, they can be defined objectively, because the criterion of ’extantness’ can always be objectively used to define a crown group: the total group can also be defined, and so the difference between them can be too. The role of the palaeontological systematist is therefore to reconstruct stem groups; i. e. to place fossil taxa in their correct stem-group and to establish their correct sister-group relationships.
The Cambrian explosion: particularly problematic, or just particularly famous?
“There is another and allied difficulty, which is much more serious. I allude to the manner in which species belonging to several of the main divisions of the animal kingdom suddenly appear in the lowest known fossiliferous rocks. ” Darwin 1859 “Taxa recognized as orders during the (Precambrian-Cambrian) transition chiefly appear without connection to an ancestral clade via a fossil intermediate. ” Valentine et al. 1991
And yet. . . ‘‘What is apparent from this restudy is that Burgessia did possess a mixture of characters. . . many of which are to be found in modern arthropods of various groups’’ (Hughes, 1975)
Trilobites are in fact an excellent example of the problem Redlichia (Hunanolenus) chinensis
“Probably no-one will now deny that trilobites are crustaceans” Raymond (1920) “The Xyphosurida [Chelicerata] are undoubtedly the closest living relatives of the Trilobita” Snodgrass (1938) “A trilobite origin for both crustaceans and chelicerates seemed most likely” Newman and Hessler (1975)
How to explain this Cambrian problem - (apparently) sudden appearance of fully-formed taxa, lots of weird animals somewhat like them?
How to explain this Cambrian problem - (apparently) sudden appearance of fully-formed taxa, lots of weird animals somewhat like them? “normal” animals here! Time? “weird” animals here! In retrospect, the solution is obvious - the Cambrian explosion is the locus classicus of the stem-/crown-group problem.
Age and stem-group length • Length of stem-group, and thus distinctness (= disparity? ) of the extant group depends on number of basal extinctions of the crown group through time: these change the concept!
Basal crown-group density and stem-group growth Dense basal crown Sparse basal crown Possible growth of stem group after single extinction (Difference being the number of in-group crown synapomorphies transferred to stem-group plesiomorphies)
Basal crown-group density and stem-group growth Dense basal crown Sparse basal crown (Difference being the number of in-group crown synapomorphies transferred to stem-group plesiomorphies)
Reason: stem groups grow from the base up (clades only evolve from their living tips!)
Stem-groups, evolution and extinction • From the previous: Stem group length depends on rate of extinction (R), not on rate of evolution: partly on the rate of extinction of the basal taxon (gappiness of the basal taxa also important). • As R = f(t) (and perhaps tends towards a Poisson distribution? ), it must be expected that stem groups steadily lengthen through time.
Growth of stem groups Budd 2003
Stem-group length and branch length • It follows that branch length (distance from basal node to living taxon) and stem group length (distance from basal node to most basal living taxon) are not the same, although branch length obviously must be > stem length). • If the rate of stem elongation is > than branch elongation, then extinction will inevitably occur. • One might think about the various relationships between stem and crown group lengths, and what they might mean!
Clade shape Long stem, short crown medium branch Long stem, long crown long branch Short stem, long crown medium branch Short stem, short crown long branch
Conclusions (I) • We expect old groups to be morphologically distinct, because they have had enough time to develop long stem-groups: e. g. the phyla • Disparity depends on extinction, not evolution!
Now consider the fossils. . . • After considering the position from the perspective of the living animals, think about trying to slot the fossils back in again. • Old groups will have long stem groups, and thus be very distinct. • The fossils that belong in their stem groups will therefore potentially be very distinct too: there is more space for them to be! • Old + distinct group = Phylum: so the relevant fossils are likely to fall into the stem groups of the phyla.
As far as the Cambrian goes… • Do we in fact find the alleged stem-group members? • Yes, but. . .
Examples of Cambrian taxa falling into the stem-groups of phyla • Echinoderms + calcichordates. Exact position controversial; but unlikely to be crown-group (e. g. ) echinoderms. • Many arthropod examples. • Palaeoscolecids etc. • Halkieria and its relatives. • Tommotiids • Hyoliths Etc etc. . .
Ctenophores Xanioascus
Annelids Burgessochaeta
Echinoderms Gogia Helicoplacoids
The problem is. . . • Identifying stem-group members is easy - but finding crown-group members is harder: 1. Crown node reconstruction for most crown groups very poor (e. g. molluscs!) 2. Need to demonstrate synapomorphy with ingroup member of crown-group - also hard.
But why only in the Cambrian? • Why do so many phyla-level stem-group forms appear in the Cambrian, but so few seem to persist later on (Devonian? ) ? • Gives the impression of “bottom-heavy” tree of life. • Obvious answer: Cambrian is time of elevated origination and extinction. Common to all radiations? • Does this intensified extinction rate affect all parts of clades, or the basal regions more severely? Might be able to detect this by examining clade density and shape.
The Cambrian explosion is stratigraphically resolvable
Can concepts be extended into the Proterozoic?
Conclusions (II) • All fossils lie in stem-groups. • Old fossils lie in deep stem-groups. • The Cambrian is packed with fossils that obviously fall into the stem-groups of phyla. • More difficult: i) detecting membership of the crown-group common to many radiations. ii) Working out why such stem-forms are apparently mostly lacking later. iii) Do stem-/crown-group distinctions tell us about the dynamics of radiations?