FIZIOLOGIJA ELIJSKE MEMBRANE nota bene Mirko A Rosic

FIZIOLOGIJA ĆELIJSKE MEMBRANE nota bene Mirko A. Rosic

CITOLOGIJA Ćelija je osnovna jedinica gradje živih bića. Osnovna supstanca ćelijskog tela je citoplazma, predstavljena mrežom lanaca posebnih proteina – polipeptida U citoplazmi – ćelijske organele (jedro, mitohondrije, endoplazmatski retikulum, ribozomi, centrozom. . . )

U jedru ćelije se može videti jedarce koje se sastoji od hromatina, posebne belanćevine koja sadrži nukleinske kiseline (genetski materijal) Centrozom ima oblik sićušne loptice (centriolus) oko koje su zrakasto grupisane vlaknaste formacije citoplazme. Igra važnu ulogu prilikom deobe ćelije Posredna deoba ćelije (mitoza), sastoji se iz 4 faze. U prvoj fazi (profaza) jedro gubi opnu i njegov hromatin obrazuje dugačka uklupčana vlakna a u centrozomu se predvoji centrioulus i njegovi delovi odlaze ka polovima ćelije dok se izmedju njih formiraju vlakanca centrosfere. U drugoj fazi (metafaza) iskidaju se hromatinska vlakna i formira se 48 hromozoma (nosioci gena). Hromozomi se rasporedjuju u dve ekvatorijalne ploče i cepaju. U trećem stadijumu (anafaza) hromozomi odlaze ka centriolusima a u četvrtoj (telofaza) oni obrazuju dva nova jedra i nastaju dve nove

HISTOLOGIJA U čovečijem telu postoje četiri osnovne vrste tkiva: epitelno, potporno, mišićno i nervno. Epitelno tkivo sastoji se samo iz ćelija koje obrazuju opne i pokrivaju spoljašnje ili unutrašnje površine tela. Ćelije epitelnog tkiva se grupišu i formiraju jednoslojne i višeslojne epitele.

Ćelije potpornog tkiva su, za razliku od epitelnog, razmaknute, postavljene na manjem ili većem rastojanju i prostore izmedju njih ispunjava medjućelijska supstanca, koja tkivu daje manju ili veću čvrstinu. Potporno tkivo se prema glavnim odlikama svoje funkcije pojavljuje u tri osnovna vida: vezivno, hrskavično i koštano. Bez potpornog tkiva bi sva ostala meka tkiva čovečijeg tela pod uticajem zemljine teže obrazovala bezobličnu pihtijastu gomilu. Vezivno tkivo može biti rastresito, fibrozno, elastično, retikularno (zvezdaste ćelije – limfni čvorovi, koštana srž, sluzokože), masno.

Hrskavično tkivo karakteriše medjućelijska sluzna belančevinasta masa, hondromukoid. U toj masi nalazi se i hondroitin-sumporna kiselina. Retke hrskavične ćelije okružene su pojedinačno ili skupno kolagenim vlaknima i obrazuju hondron – osnovni funkcionalni elemenat gradje hrskavice. Izmedju hondrona pružaju se kolagena vlakna ka površini hrskavice i spajaju se sa vezivnim vlaknima opne hrskavice - perihondrijumom. Koštane ćelije (osteociti) imaju izdužen, elipsoidni oblik, s mnogobrojnim tankim nastavcima, koji se dodiruju. Osnovna medjućelijska masa sastoji se iz snopova kolagenih vlakana duž kojih se u vidu sitnih kristalića talože mineralne soli i to uglavnom kalcijum fosfat (85%) i kalcijum karbonat (10%) a u manjoj meri magnezijum fosfat, kalcijum fluorid, i kalcijum hlorid. Kolagena vlakna i koštane ćelije predstavljaju organski deo koštanog tkiva – osein (30%) Osnovna supstanca koštanog tkiva nije homogena već u slojevima – lamele. Lamele imaju izgled šupljih valjkova koji se uvlače jedan u drugi

Mišićno tkovo je izgradjeno iz ćelija koje su sposobne da se pod uticajem nadražaja kontrahuju, skraćuju. U čovečijem telu se razlikuju tri vrste mišićnog tkiva: - poprečnoprugasto - glatko - srčano

Organizacija skeletnog mišića od makroskopskog do molekulskog nivoa. F, G, H, I su poprečni preseci na označenom nivoima. Područje između dva “Z” diska naziva se sarkomera i predstavlja osnovnu funkcionalnu jedinicu.
 Nervna ćelija je najdiferenciranija ćelija")
Nervno tkivo sačinjavaju nervne ćelije i potporne ćelije (neuroglije) Nervna ćelija je najdiferenciranija ćelija u našem telu a njena uloga je da prima i sprovodi nadražaje.

Organ je deo čovečijeg tela odredjenog oblika i funkcije. On se sastoji iz više tkiva, od kojih je jedno dominantno, glavni nosilac funkcije karakteristične za dotični organ. Ostala pomoćna tkiva imaju pomoćnu ulogu u funkciji organa. Organi se udružuju i grade pribore ili sisteme čovečijeg tela. Organi jednog sistema iako imaju različite uloge, ipak sinergično omogućavaju izvodjenje jedne složene funkcije čovečijeg organizma. U čovečijem telu postoje sledeći funkcionalni sistemi: - skeletni sistem - mišićni sistem - pribor za varenje (digestivni sistem) - pribor za disanje (respiratorni sistem) - mokraćno-spolni pribor (genitourinarni sistem) - pribor za krvotok (kardiovaskularni sistem) - nervni sistem - sistem čula - koža

ŠTA STVARA I ODRŽAVA RAZLIKU U SASTAVU EKSTRAĆELIJSKE I INTRAĆELIJSKE TEČNOSTI ? ? ?

ĆELIJSKA MEMBRANA !!!

LIPIDNI DVOSLOJ - Fosofolipidi sadrže glicerolske glave koje su hidrofilne i dva repa sastavljena od masnih kiselina koji su hidrofobni. Hidrofobni repovi orijentišu se jedni naspram drugih i formiraju dvosloj. - Liposolubilne supstance (kiseonik, ugljendioksid steroidni hormoni…) lako prolaze kroz ćelijsku membranu jer se lako rastvaraju u hidrofobnom lipidnom dvosloju. - Hidrosolubilne supstance (joni natrijuma ili hlora, glukoza…) nemogu se rastvoriti u lipidnom dvosloju membrane, ali mogu proći kroz nju koristeći vodom ispunjene kanale ili pore, ili mogu biti transportovane pomoću specijalnih nosača.

Proteini membrane - Integralni proteini -prolaze kroz celu membranu -igraju ulogu jonskih kanala ili transportnih proteina (nosača) - Periferni proteini -nalaze se ili sa unutrašnje ili sa spoljašnje strane membrane -igraju ulogu receptora u procesu prenošenja informacija sa ćelije na ćeliju
 - Gap junctions")
MEDJUĆELIJSKE KONEKCIJE - Tight junctions (zonula occludens) - Gap junctions
 -najčešće izmedju epitelnih ćelija -mogu biti medjućelijski putevi za rastvotrne")
Tight junction (čvrste veze) -najčešće izmedju epitelnih ćelija -mogu biti medjućelijski putevi za rastvotrne čestice, zavisno od nihove veličine, električnog naboja, kao i od karakteristika same veze. -mogu biti nepropusne, kao kod distalnih renalnih tubula ili propusne (“leaky”) kao kod proksimalnih renalni tubula i žučne kesice
 -spojevi izmedju ćelija koji dozvoljavaju medjućelijsku komunikaciju -dozvoljavaju napr. prenošenje")
Gap junction (porozne veze) -spojevi izmedju ćelija koji dozvoljavaju medjućelijsku komunikaciju -dozvoljavaju napr. prenošenje električnih potencijala sa jedne na drugu srčanu ćeliju ili u hemijskim sinapsama.
 -posredovan nosačima (olakšana difuzija, primarni i")
TRANSPORT KROZ ĆELIJSKU MEMBRANU -difuzija (prosta i olakšana) -posredovan nosačima (olakšana difuzija, primarni i sekundarni aktivni transport)

DIFUZIJA – svojstvo svih čestica u telesnim tečnostima da se haotično kreću predajući deo svoje kinetičke energije okolnim česticama. (fiz. =toplota; što je kretanje brže – temperatura je viša) Kretanje čestica prestaje samo na temperaturi apsolutne nule BIOLOŠKI ZAKONI (PRAVILA)

EFEKAT KONCENTRACIJSKOG, ELEKTROHEMIJSKOG GRADIJENTA I PRITISKA NA DIFUZIJU ČESTICA KROZ ĆELIJSKU MEMBRANU
 lako prolaze kroz ćelijsku membranu jer se")
- Liposolubilne supstance (kiseonik, ugljendioksid steroidni hormoni…) lako prolaze kroz ćelijsku membranu jer se lako rastvaraju u hidrofobnom lipidnom dvosloju. - Hidrosolubilne supstance (joni natrijuma ili hlora, glukoza…) nemogu se rastvoriti u lipidnom dvosloju membrane, ali mogu proći kroz nju koristeći vodom ispunjene kanale ili pore, ili mogu biti transportovane pomoću specijalnih nosača.

PROSTA DIFUZIJA -jedini oblik transporta koji nije posredovan nosačem -dogadja se u pravcu elektrohemijskog gradijenta (“nizbrdo”) -ne zahteva energiju – pasivan transport -izračunava se na osnovu sledeće formule: gde je: J=-PA(C 1 - C 2) J=fluks ili protok čestica mmol/sec P=permeabilnost (cm/sec) A=površina polja na kome se odvija difuzija (cm 2) C=koncentracija (mmol/L) * Znak minus se može ignorisati ako je C 1>C 2
: -”P” iz jednačine za difuziju J=-PA(C 1 - C 2) -opisuje lakoću")
PERMEABILNOST (propustljivost): -”P” iz jednačine za difuziju J=-PA(C 1 - C 2) -opisuje lakoću kojom rastvorena čestica prolazi kroz membranu -zavisi od karakteristika* rastvorene čestice i od karakteristika membrane * probaj da se setiš nekih karakteristika !!!

-Faktori koji povećavaju permeabilnost: -↑ ulje/voda particioni koeficijent = veća rastvorljivost u lipidnom dvosloju -↓ veličina rastvorene supstance = veća brzina difuzije -↓ debljina membrane = smanjena distanca kretanja -Male hidrofobne čestice imaju najveću permeabilnost - Hidrofilne čestice prolaze ćelijsku membranu kroz vodom ispinjene kanale ili pore - Ako je čestica naelektrisana (jon), onda će njen prolaz kroz membranu zavisiti I od koncentracijskog gradijenta i od razlike u električnom potencijalu ćelijske membrane + - -
: -Stereospecifičnost. - pravi razliku")
NOSAČIMA POSREDOVAN TRANSPORT (olakšana difuzija, primarni i sekundarni aktivni transport): -Stereospecifičnost. - pravi razliku izmedju dva izomera (prosta difuzija ne) D-glukoza se transportuje olakšanom difuzijom; njen L-izomer nemože. -Saturabilnost. – brzina transporta raste dok se nosačI ne zasite. Transportni maksimum (Tm) odgovara maksimalnoj brzini (Vmax) u enzimskoj kinetici -Kompetitivnost. – strukturno slične čestice konkurišu jedna drugoj za transportna mesta na nosačkom molekulu (galaktoza je kompetitivni inhibitor transporta glukoze u intestinumu)
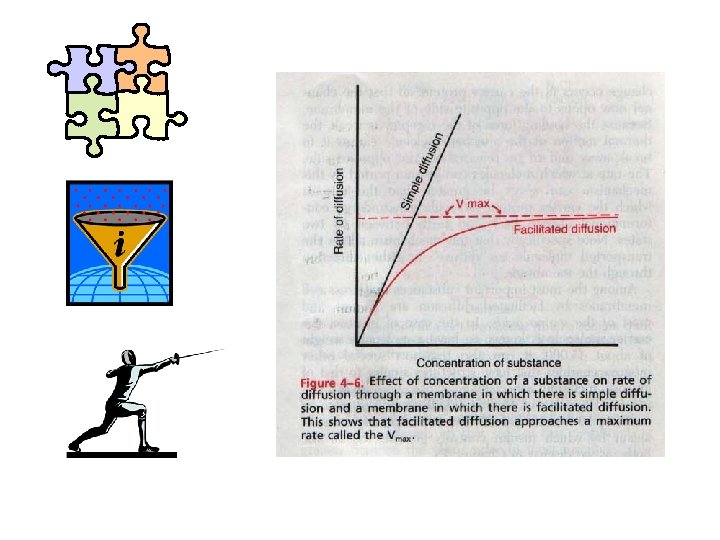
, slično kao i prosta difuzija.")
OLAKŠANA DIFUZIJA: -odvija se u pravcu elektrohemijskog gradijenta (nizbrdo), slično kao i prosta difuzija. -ne zahteva utrošak metaboličke energije i zato je to pasivan (a ne aktivan) transport. -mnogo je brža u poredjenju sa prostom difuzijom. -spada u transporte posredovane nosačem i zato važe pravila stereospecifičnosti saturabilnosti i kompetitivnosti

PRIMER OLAKŠANE DIFUZIJE: -Transport glukoze u mišićnim i ćelijama masnog tkiva: -odvija se “nizbrdo” -posredovan je nosačem -inhibira ga galaktoza Proteinski nosači koji učestvuju u olakšanoj difuziji glukoze zahtevaju prisustvo insulina !!!
 -zahteva utrošak metaboličke energije u")
PRIMARNO AKTIVNI TRANSPORT: -odvija se suprotno elektrohemiskom gradijentu (“uzbrdo”) -zahteva utrošak metaboličke energije u formi ATP-a i zato se naziva aktivnim -posredovan je nosačem pa zato važe pravila stereospecifičnosti, kompetitivnosti i saturabilnosti
 održava nisku intraćelijsku konventraciju")
PRIMERI PRIMARNO AKTIVNOG TRANSPORTA -Na+, K+- ATPasa (Na+, K+- pumpa) održava nisku intraćelijsku konventraciju Na+, i visoku intraćelijsku koncentraciju K+. Oba jona se transportuju suprotno njihovom elektrohemijskom gradijentu. Energija se obezbedjuje iz terminalnih P veza ATP-a. Joni se transportuju u odnosu = 3 Na+/2 K+. Specifični inhibitori su glikozidi ouabain i digitalis. -Ca 2+-ATPasa (Ca 2+-pumpa) na sarkoplazmatskom retikulumu ili na ćelijskim membranama transportuje jon kalcijuma suprotno elektrohemijskom gradijentu -H+, K+- ATPasa (ili protonska pumpa) u gastričnim parijetalnim ćelijama, transportuje protone u lumen želuca Suprotno elektrohemijskom gradijentu a inhibira se Omeprazolom.

SEKUNDARNO AKTIVNI TRANSPORT: -transport dve ili više čestica je povezan -jedna od čestica (obično Na+) transportuje se “nizbrdo” I obezbedjuje energiju za “uzbrdo” transport druge -energija se ne obezbedjuje direktno već indirekto iz gradijenta za Na+ koji se stalno održava. (inhibicija Na+, K+- ATPase može zaustaviti sekundarno aktivni transport) -ako se čestice kreću u istom pravcu kroz membranu to se naziva kotransport ili simport (Na+-glucosa kotransport u crevima ili Na+-K+-2 Cl+ kotransport u tankom ascedentnom delu henleove petlje u bubrezima. -Ako se čestice kreću u suprotnim pravcima kroz membranu to se naziva kontratransport, razmena ili antiport (Na+-Ca 2+ i Na+-H 2+ - razmena)

Na+-glukoza kotransport -nosač se nalazi na luminalnoj strani membrane u intestinalnoj mukozi irenalnim proksimalnim tubulima -glukoza se transportuje “uzbrdo” dok se natrijum transportuje “nizbrdo” -gradijent za natrijum održava se aktivnošću Na+, K+- pumpe na bazolateralnoj strani membrane. Inhibicija pumpe smanjuje transmembranski gradijent za natrijum I indirektno inhibira ovaj transport.

Na+-Ca 2+ kontratransport ili razmena -transport Ca 2+ “uzbrdo iz niske intraćeliske u visoku ekstraćelijsku koncentraciju Ca 2+ dok se Na+ kreće u suprotnom pravsu kroz ćelijsku membranu - gradijent za natrijum održava se aktivnošću Na+, K+- pumpe. Inhibicija pumpe smanjuje transmembranski gradijent za natrijum i indirektno smanjje Na+-Ca 2+ razmenu.

KARAKTERISTIKE RAZLIČITIH TIPOVA TRANSPORTA Tip Elektrohemijski gradijent Posredovan nosačem Metabolička energija Na+ gradient inhibicija Na+K+ pumpe Prosta difuzija Nizbrdo Ne Ne Ne - OlakŠana difuzija Nizbrdo Da Ne Ne - Primarno aktivni Uzbrdo Da Da - Kotransport uzbrdo * Da Indirektno Da u istom pravcu Kontratransportt uzbrdo * da indirektno Da u suprotnom inhibira pravcu * Jedna ili više čestica transportuje se uzbrdo; Na+ se transportuje nizbrdo Inhibira
 kroz polupropustljivu ćelijsku membranu, iz rastvora sa malom")
OSMOZA je protok vode (neto difuzija) kroz polupropustljivu ćelijsku membranu, iz rastvora sa malom koncentracijom rastvorenih čestica u rastvor sa većom koncentracijom rastvorenih čestica!!!

-Rastvori A I B razdvojeni su semipermeabilnom membranom. Rastvor A sadrži čestice koje su isuviše velike da bi prošle kroz membranu. Rastvor B je čista voda. Prisustvo čestica u rastvoru A dovodi do nastanka osmotskog pritiska -Razlika u osmotskim pritiscima koja se razvija na membrani uzrokuje da se voda kreće iz rastvora B (koji ima niži osmotski pritisak) u rastvor A ( sa višim osmotskim pritiskom) -Vremenom će se volumen rastvora A povećati a volumen rastvora B smanjiti.
 u rastvoru naziva se OSMOLARNOST -Osmolarnost se može")
-Koncentracija osmotski aktivnih čestica (nedifusibilne čestice) u rastvoru naziva se OSMOLARNOST -Osmolarnost se može izračunati korišćenjem sledeće formule: O=gx. C gde je: O = osmolarnost (osm/L) g = broj čestica u rastvoru (osm/mol) C = koncentracija (mol/L) g. Na. Cl = 2 gglucose = 1 Na. Cl ↔ Na+ + Cl-

ISOSMOTSKI = rastvori iste osmolarnosti HIPEROSMOTSKI = rastvor sa većom osmolarnošću HIPOOSMOTSKI =rastvor sa nižom osmolarnošću

OSMOTSKI PRITISAK se može izračunati na osnovu Van’t Hoff’s –ovog zakona koji tvrdi da osmotski pritisak zavisi od koncentracije osmotski aktivnih čestica. Koncentracija čestica konvertuje se u pritisak na osnovu sledeće formule: Π = g x C x RT gde je: - Π = osmotski pritisak (mm. Hg) - g = broj čestica u rastvoru (osm/mol) - C = koncentracija (mol/L) - R = gasna konstanta - T = absolutna temperatura

-Osmotski pritisak raste kada koncentracija rastvora raste - Što veći osmotski pritisak rastvora to veći priliv vode - IZOTONIČNI = rastvori sa istim efektivnim osmotskim pritiskom - HIPERTONIČAN = rastvor sa većim efektivnim osmotskim pritiskom - HIPOTONIČAN = rastvor sa nižim efektivnim osmotskim pritiskom -Koloidno osmotski ili onkotski pritisak je osmotski pritisak koga proizvode proteini (npr. proteini plazme) - Uporedi (osmolarnost i osmotski pritisak): 1 M Ca. Cl 2 1 M KCl O=gx. C Π = g x C x RT SETI SE !!!


JONSKI KANALI su integralni proteini koji se prostiru kroz celu membranu, od njene spoljašnje do unutrašnje strane, i kada su otvoreni omogućavaju prolazak određenih jona.

KARAKTERISTIKE JONSKIH KANALA: - Jonski kanali su selektivni; omogućavaju prolazak samo određenih jona. Selektivnost se zasniva na veličini kanala i rasporedu naelektrisanja unutar njih. (Mali kanal koji je ispunjen negativno naelektrisanim grupama će tako biti selektivan za male katjone i neprolazan za velike molekule i anjone) - Jonski kanali mogu biti otvoreni ili zatvoreni. Kada je kanal otvoren tada joni za koje je selektivan mogu da prođu kroz njega. Kada je kanal zatvoren, joni ne prolaze kroz njega.

OTVARANJE I ZATVARANJE KANALA JE KONTROLISANO VRATIMA -Voltaž zavisni kanali se otvaraju i zatvaraju u zavisnosti od promena membranskog potencijala - Ligand zavisni kanali su otvoreni ili zatvoreni u zavisnosti od prisustva specifičnog molekula: hormona, drugih glasnika ili neurotransmitera.

PRIMERI VOLTAŽ ZAVISNIH KANALA -Aktivaciona vrata Na+ kanala u nervu se otvaraju depolarizacijom i kada su otvorena, membrana nerva je propustljiva za Na+. -Inaktivaciona vrata Na+ kanala se zatvaraju depolarizacijom; kada su zatvorena, membrana je nepropustljiva za Na+. -Vrata (samo jedna) za K+ se otvaraju depolarizacijom; kada su otvorena membrana je propustljiva za K+. !!!

PRIMER ZA LIGAND ZAVISNE KANALE -Nikotinski receptor za acetilholin na motornoj završnoj u mišićima ploči je jonski kanal koji se otvara kada se acetilholin veže za njega. Kada je otvoren, propustljiv je za Na+ i K+, što dovodi do depolarizacije motorne završne ploče.

DIFUZIONI I RAVNOTEŽNI POTENCIJALI -Difuzioni potencijal je razlika potencijala duž membrane koja nastaje kao posledica jonske koncentracione razlike i difuzije jona kroz membranu. -Difuzioni potencijal može da nastane samo ako je membrana propustljiva za jon. -Vrednost difuzionog potencijala zavisi od veličine koncentracionog gradijenta. -Znak difuzionog potencijala zavisi od toga da li je jon koji difunduje pozitivno ili negativno naelektrisan. -Difuzioni potencijali su nastali difuzijom malog broja jona i ne dovode do promena u koncentraciji jona koji difunduju. -Ravnotežni potencijal je difuzioni potencijal koji tačno uravnotežava, suprotstavlja se, tendenciji za difuzijom, a tendencija za difuzijom nastaje kao posledica razlike u koncentraciji jona. Pri elektrohemijskoj ravnoteži, sile hemijskog i električnog kretanja nekog jona su jednake i suprotne, tako da neto difuzija jona više ne postoji. + + - - -
 sile električnog")
PRIMERI ZA Na+ I K+ DIFUZIONE POTENCIJALE sile hemijskog kretanja (koncentraciona razlika) sile električnog kretanja (potencijalna razlika)

NERNST-ova JEDNAČINA se koristi za izračunavanje ravnotežnog potencijala za datu koncentracionu razliku permeabilnog jona kroz ćelijsku membranu. E = ± 61 log Ci/Co gde je Co = ekstracelularna koncentracija i Ci = intracelularna koncentracija E Na+= + 65 m. V EK+= - 85 m. V ECa 2+ = + 120 m. V ECl- = - 85 m. V Ova jednačina pokazuje vrednost potencijala za dati jon pri elektrohemijskoj ravnoteži u slučaju da je membrana permeabilna samo za taj jon (ne i za druge jone) Znak potencijala ima pozitivnu vrednost (+) ako je jon koji se razmatra negativan a negativnu (-) ako je jon pozitivan. Izračunajte membranski potencijal za različite vrednosti C i razmislite!!!

GOLDMAN-HODKIN-KATZ-ova JEDNAČINA -Kada je membrana permeabilna za nekoliko različitih jona, difuzioni potencijal koji se razvija zavisi od : 1. polarnosti električnog naboja za svaki jon 2. permeabiliteta membrane za svaki jon (P) 3. koncentracije (C) posmatranih jona sa unutrašnje (i) ili spoljašnje (o) strane membrane. PNa i PK se brzo menja tokom provođenja impulsa (u ekscitabilnim tkivima) Izračunaj membranski potencijal za različite vrednosti C i P (od 0 do 1) i razmisli!!!

MIROVNI MEMBRANSKI POTENCIJAL -se izražava kao izmerena potencijalna razlika kroz ćelijsku membranu u m. V. -se prema konvenciji, izražava kao vrednost intracelularnog potencijala u odnosu na ekstracelularni potencijal. Tako, mirovni membranski potencijal od – 70 m. V znači 70 m. V, sa negativnošću u ćeliji.
 nastaje kao posledica difuzionog potencijala koji sa druge strane nastaje")
-Mirovni membranski potencijal (MMP) nastaje kao posledica difuzionog potencijala koji sa druge strane nastaje kao rezultat koncentracione razlike permeabilnih jona. - Svaki permeabilni jon teži da upravi membranski potencijal prema svom ravnotežnom potencijalu. Joni koji poseduju najveću permeabilnost ili konduktansu, će imati najveće učešće u formiranju MMP. -Tokom mirovanja, nervna membrana je daleko više permeabilna za K+ nego Na+. Zbog toga je vrednost MMP blizu vrednosti izračunatog ravnotežnog potencijala za K+, ali daleko od Na+ ravnotežnog potencijala. -Direktno učešće elektrogene Na+-K+ pumpe je malo. Pumpa učestvuje u stvaranju MMP više indirektno, na taj način što održava gradijent koncentracije kroz ćelijsku membranu
 koje se sastoji iz")
AKCIONI POTENCIJAL - je svojstvo ekscitabilnih ćelija (tj. nerva, mišića) koje se sastoji iz brze (msec) depolarizacije, koja je praćena repolarizacijom membranskog potencijala. Akcioni potencijali imaju: - stereotipičnu veličinu i oblik - prostiru se (šire se) duž membrane - po principu sve ili ništa.
. HIPERPOLARIZACIJA povećava negativnost membranskog")
DEPOLARIZACIJA smanjuje negativnost membranskog potencijala (unutrašnjost ćelije postaje manje negativna). HIPERPOLARIZACIJA povećava negativnost membranskog potencijala (unutrašnjost ćelije postaje više negativna). "UNUTRAŠNJA STRUJA" (INWARD CURRENT) je kretanje pozitivnog naelektrisanja u ćeliju. “Unutrašnja struja “ depolarizuje membranu "SPOLJNA STRUJA" OUTWARD CURRENT je kretanje pozitivnog naelektrisanja iz ćelije. "Spoljna struja" hiperpolarizuje membranu.

PRAG je vrednost potencijala membrane na kome je akcioni potencijal neizbežan. Inward current (unutrašnja struja) depolarizuje membranu. Ako unutrašnja struja depolarizuje membranu do vrednosti praga, nastaje akcioni potencijal. U slučaju da nije dovoljno pozitivnih jona ušlo u ćeliju da bi depolarizovalo membranu do praga (subpragovni potencijal), tada neće nastati akcioni potencijal.

AKCIONI POTENCIJAL - Kretanje pozitivnih jona u ćeliju depolarizuje membranski potencijal do praga - Depolarizacija prouzrokuje brzo otvaranje aktivacionih vrata Na+ kanala, i provodljivost za Na+ brzo raste. - Provodljivost za Na+ postaje veća nego za K+ , i membranski potencijal se kreće prema (ali ga sasvim ne dostiže) ravnotežnom potencijalu za Na+. Takva brza depolarizacija tokom akcionog potencijala nastaje usled ulaska Na+ u ćeliju. -The overshot je brzi deo na vrhu akcionog potencijala kada je potencijal membrane pozitivan. -Tetrodotoxin (TTX) blokira ove voltaž-zavisne Na kanale i onemogućava nastajanje akcionog potencijala.

FAZA REPOLARIZACIJE AKCIONOG POTENCIJALA -Depolarizacija izaziva i zatvaranje inaktivacionih vrata Na+ kanala (ali mnogo sporije nego što uzrokuje otvaranje aktivacionih vrata). Zatvaranje inaktivacionih vrata dovodi do zatvaranja natrijumovih kanala, i provodljivost za Na+ se vraća ka nuli. -Depolarizacija sporo otvara K+ kanale i povećava provodljivost za K+, čak do višeg nivoa nego u mirovanju - Zajednički efekat zatvaranja natrijumovih kanala i veće otvaranje kalijumovih kanala dovodi do povećanja provodljivosti za kalijum, i membranski potencijal je repolarizovan. Na taj način, izlazak kalijumovih jona iz ćelije dovodi do repolarizacije. -Undershoot (hyperpolarizing afterpotential or positive afterpotential) hiperpolarizujući naknadni potencijal ili pozitivni naknadni potencijal – Provodljivost za kalijum se zadržava na višoj vrednosti nego u mirovanju kratko vreme nakon zatvaranja natrijumovih kanala. Tokom ovog perioda, membranski potencijal je veoma blizu vrednosti ravnotežnog potencijala za kalijum.

REFRAKTORNI PERIODI: - Apsolutni refraktorni period - Relativni refraktorni period

Apsolutni refraktorni period je period tokom koga drugi akcioni potencijal ne može da se izazove bez obzira koliko jak bio stimulus. Ovaj period se podudara sa celim vremenom trajanja akcionog potencijala (najmanje za 2/3 trajanja akcionog potencijala). Prisetite se da su inaktivaciona vrata Na+ kanala zatvorena tokom depolarizacije membrane. Ona ostaju zatvorena sve dok traje proces repolarizacije. Akcioni potencijal ne može da nastane sve dok su inaktivaciona vrata zatvorena.

Relativni refraktorni period počinje na kraju apsolutnog refraktornog perioda i nastavlja se sve dok se membranski potencijal ne vrati do mirovnog nivoa. Akcioni potencijal može da se izazove tokom ovog perioda jedino u slučaju ako je povećan (više nego obično) ulazak pozitivnih jona u ćeliju. Provodljivost za K+ je veća nego u mirovanju, membranski potencijal je blizu ravnotežnog potencijala za K+ i, zbog toga, udaljeniji od praga; za dostizanje praga potreban je veći ulazak pozitivnih jona u ćeliju.

AKOMODACIJA se javlja kada se vrednost potencijala ćelijske membrane održava na depolarizacionom nivou tako da pragovni potencijal postoji ali bez okidanja akcionog potencijala. Akomodacija može da se demonstrira preko hiperkalemije, kada je skeletna mišićna membrana depolarizovana usled visoke koncentracije ekstracelularnog K+. Iako je membranski potencijal blizu praga, akcioni potencijal se ne javlja jer su inaktivaciona vrata Na+ kanala zatvorena usled depolarizacije, što dovodi do mišićne slabosti.

Propagacija akcionih potencijala nastaje kada se lokalno kretanje pozitivnih jona proširi na susedne delove membrane, koji se zatim depolarizuju do pragovnog potencijala nakon čega nastaju akcioni potencijali.

Brzina provođenja u nervnim vlaknima zavisi od: a. Dijametra vlakna – povećanje dijametra nervnog vlakna dovodi do smanjenja unutrašnjeg otpora; tako, brzina provođenja impulsa raste što je vlakno deblje. b. Mijelinske ovojnice. Mijelin ima ulogu izolatora oko aksona i povećava brzinu provođenja impulsa u nervnim vlaknima. Mijelinizirana nervna vlakna provode impulse tzv. saltatornom kondukcijom budući da akcioni potencijali mogu da nastanu samo u Ranvijerovim čvorovima, mestima prekida mijelinske ovojnice.
- Slides: 63