Fatores que controlam a atividade enzimtica 3 Concentrao






 é um parâmetro cinético que traz informações sobre a")

![A velocidade da reação somente é proporcional à [E] quando a enzima está saturada,](https://slidetodoc.com/presentation_image_h2/559d104e4ff8702f52a4a0f47da5cc47/image-10.jpg "A velocidade da reação somente é proporcional à [E] quando a enzima está saturada,")
 Substratos")


![û Gráfico de Eadie-Hofstee Vmax v Inclinação = -Km Vmax Km v [S]](https://slidetodoc.com/presentation_image_h2/559d104e4ff8702f52a4a0f47da5cc47/image-14.jpg "û Gráfico de Eadie-Hofstee Vmax v Inclinação = -Km Vmax Km v [S]")
![û Gráfico de Hanes-Woolf [S] v Inclinação = 1 Vmax Km Vmax -Km [S]](https://slidetodoc.com/presentation_image_h2/559d104e4ff8702f52a4a0f47da5cc47/image-15.jpg "û Gráfico de Hanes-Woolf [S] v Inclinação = 1 Vmax Km Vmax -Km [S]")



- Slides: 18
Fatores que controlam a atividade enzimática: 3. Concentração: • da enzima • do substrato • de co-fatore(s) A [substrato] cai na mesma razão em que a [produto] aumenta em função do tempo. A enzima existe sob duas formas: enzima livre E e complexo enzima-substrato ES. No início da reação, a [E] livre cai e a do complexo [ES] aumenta e atinge um máximo, em que não há mais [E] livre no meio. Nessa situação (indicada no retângulo cinza), diz-se que a enzima está saturada (só existe no complexo ES). A velocidade da reação é a máxima. O gráfico abaixo ilustra como as concentrações de E, S e P variam ao longo do tempo da reação. concentração 2. Tempo da reação tempo
Influencia da quantidade de enzima na velocidade de conversão vo Este fato permite entender que se use a atividade enzimática como uma medida da quantidade de enzima
Modelos empíricos para correlacionar a hidrólise enzimática com o tempo de reação Com os dados de rendimento de hidrolise para cada carga de enzima e tempo pode-se fazer a modelagem -Há uma relaçao logaritmica linear entre o rendimento de hidrolise e o tempo? - Há uma relaçao logaritmica linear entre o rendimento de hidrolise e a carga de enzima? -Há uma relaçao logaritmica linear entre a variação do rendimento de hidrolise (coeficiente angular ) e o tempo?
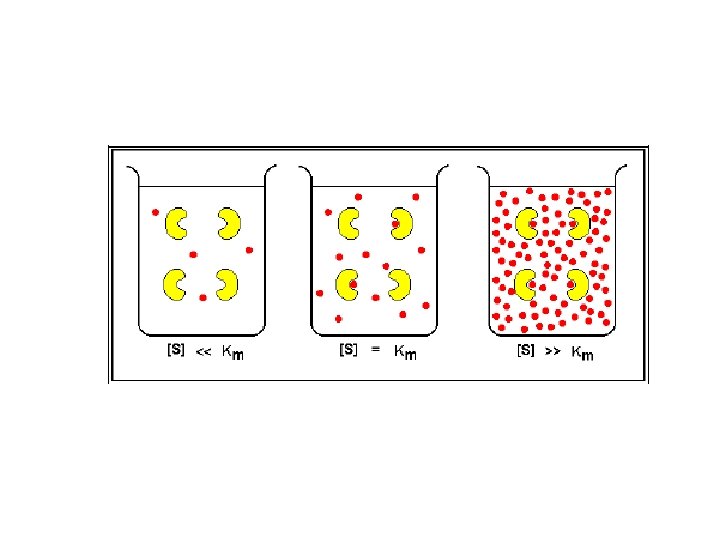
Influencia da concentração de substrato na velocidade de conversão vo A velocidade da reação apresenta três regiões de comportamento diferente, a medida que se aumenta a concentração do substrato -parte a: v aumenta proporcionalmente com aumentos de S. -parte b: v aumenta não proporcionalmente com aumentos de S. -parte c: v não aumenta mais, tendendo a um valor máximo (Vmax), sendo independente da [S]
Na década de 1950: Michaelis e Menten formularam as bases da cinética enzimática, para explicar como a concentração do substrato [S] afeta a velocidade da reação v, conforme se observa no gráfico abaixo. : O gráfico mostra um conjunto de reações que estão acontecendo simultaneamente, conforme as equações abaixo:
Para se chegar à equação da hipérbole quadrada do gráfico V x S, o gráfico de Michaelis Menten, considera-se que o conjunto de reações está em equilíbrio, ou seja, a [ES] é constante e o sistema tem a sua velocidade máxima, Vmax. Quando [ES] é constante, as velocidades de formação (Vf) e de desdobramento (Vd) do complexo ES são iguais: Vf formação ES = Vd desdobramento ES Aplicando a Lei de Ação das Massas para definir Vf e Vd, temos: Vf = k 1 [ES] [E]+[S] Vd= k-1 [E]+[S] + k 2 [E]+[P] [ES] Igualando-se Vf = Vd e resolvendo para V, chega-se à Equação de Michaelis-Menten: sendo KM = k-1+ k 2 k 1 Constante de Michaelis-Menten
A constante de Michaelis-Menten (KM) é um parâmetro cinético que traz informações sobre a afinidade que a enzima tem pelo substrato. O KM é numéricamente igual à [substrato] que produz metade da Vmax. Substituindo na equação v por Vmax/2, vemos: Vo=Vmax = Vmax. [S] 2 2 Km + [S] Vmax. Km + Vmax. [S] = 2 Vmax. [S] KM = k-1+ k 2 Vmax. Km = Vmax. [S] k 1 Km = [S] Considerando a afinidade da E pelo seu S, temos 2 casos: 1. E tem baixa afinidade por S Quando Vd é mais alta do que Vf k-1+ k 2 = grande k 1 = pequeno Km alto 2. E tem alta afinidade por S Quando Vf é mais alta do que Vd k-1+ k 2 = pequeno k 1 = grande Km baixo
Uma outra forma de se obter os valores de KM e de Vmax é através do gráfico dos duplos recíprocos (1/V x 1/S), e da equação de Lineweaver-Burk. Aplicando-se o inverso a ambos os lados da equação de Michaelis-Mentem, obtem-se a equação de Lineweaver-Burk, que é uma função linear (uma reta): Tang a =KM/Vmax O intercepto da reta no eixo x é igual a -1/KM substituindo y = 0 na equação, temos: 0 = Km. 1 + 1 Vmax [S] Vmax -1 = Km [S] -1 = Km. 1 Vmax [S] 1 [S] y = a. x + b equação de reta Onde: a = coef. angular = Km/Vmax (tangente ângulo alfa) b = coef. linear = 1/Vmax (intercepto no eixo y)
A velocidade da reação somente é proporcional à [E] quando a enzima está saturada, ou seja, reação é de ordem zero (independe) em relação a [S] Eficiência catalítica Kcat/Km - k 2 ou kcat (constante catalítica) mede o “poder catalítico” da enzima v = k 2 Etotal + [P] [ES] k 2 = Vmax (s-1) [Etotal] Parâmetro mais adequado para comparações cinéticas Para calcular kcat considera-se que toda a E existe como ES, e que v=Vmax
Interações secundárias de proteinases com seus substratos Enzima Tripsina (p. H 7. 5) Substratos Z-Lys-OMe Z-(Ala)2 -Tyr-Lys-OMe kcat (seg-1) KM (m. M) Razão kcat/ KM 101 106 0. 23 0. 08 1 3 Quimotripsina (p. H 7. 9) Z-Tyr-Gly-OMe Z-(Ala)2 -Ala-OMe 0. 6 10 23 2 1 190 Elastase (p. H 8. 0) Z-Ala-OMe Z-(Ala)2 -Ala-OMe 6. 7 73 153 0. 43 1 3900 A tabela ilustra como interpretar Kcat, KM e eficiência catalítica (kcat/KM), comparando a ação de enzimas proteolíticas sobre diferentes substratos sintéticos. O ponto de clivagem dos substratos está indicado pela flecha. Conforme o número de resíduos de aminoácidos aumenta à esquerda do ponto de clivagem, a eficiência catalítica (kcat/KM) das enzimas melhora (considerando-se como 1 o valor obtido com o substrato mais curto). No caso da tripsina, o Km diminue 3 X, sem alterar o kcat, ou seja a formação do complexo ES com o substrato maior é facilitado, mas não foi afetada a sua transformação em produto. No caso da elastase, o Km diminue ~300 X e o kcat aumenta ~10 x, ou seja todas as etapas da reação são facilitadas com o substrato mais longo. Como resultado, a eficiência catalítica aumentou 3. 900 vezes.
35Å QUIMOTRIPSINA 18 Å SÍTIO ATIVO SUBSTRATO S 4 S 3 S 2 S 1¹ P 4 P 3 P 2 P 1¹ SÍTIO CATALÍTICO LIGAÇÃO SUSCEPTÍVEL Representação esquemática do sítio de reconhecimento do substrato da QUIMOTRIPSINA. O sítio ativo de uma enzima proteolítica em geral é uma fenda onde se localizam os resíduos de aminoácidos (S) dotados de capacidade catalítica. O resíduo de aminoácido da enzima que efetua a clivagem do substrato é designado S 1, e sucessivamente à esquerda na seqüência primária da enzima, ficam os resíduos S 2, S 3, S 4, etc. De maneira análoga, os resíduos de aminoácidos da enzima à direita de S 1 são designados com S’ 1, S’ 2, S’ 3, etc. O resíduo catalítico S 1 da enzima cliva o substrato proteico no resíduo P 1, determinando sua especificidade primária. À esquerda e à direita de P 1 na seqüência do substrato proteico, estão os resíduos P 2, P 3, etc, e P’ 1, P’ 2, etc, respectivamente. A figura acima explica os dados da tabela no slide anterior. O encaixe da enzima com a molécula do substrato melhora quando outros resíduos, além de S 1 e P 1, interagem. Na tabela, os substratos com resíduos P 2 a P 4 proporcionaram melhores parâmetros, mostrando que as interações que esses realizam com a enzima influenciam sua atividade enzimática.
MÉTODOS GRÁFICOS û Gráfico dos Recíprocos de Lineweaver-Burk 1 v Inclinação = 1 Vmax -1 Km 1 [S] Km Vmax
û Gráfico de Eadie-Hofstee Vmax v Inclinação = -Km Vmax Km v [S]
û Gráfico de Hanes-Woolf [S] v Inclinação = 1 Vmax Km Vmax -Km [S]
Inibição competitiva I S S S P S SS IS E S I S S • o inibidor é análogo estrutural do substrato e compete com ele pela ligação ao sítio ativo • com aumento da [substrato], ocorre diminue da inibição caracterizando uma competição entre S e I • não há alteração da Vmax • há um aumento de Km por um fator a, que permite o cálculo da constante de inibição, Ki
I S Inibição não competitiva ou mista S • inibidor não é análogo estutural do E S P I I I S I E E I S S S I I S S substrato - não se liga ao sítio ativo • inibidor se liga à E e ao ES • aumento da [substrato] não diminue a inibição - não há competição • Km aumenta e Vmax diminuí
E S I I Inibição incompetitiva S S • inibidor é análodo do estado de transição e se liga somente ao complexo ES • Km aumenta e Vmax diminuí