EKOFIZIOLOKE ODLIKE C4 i CAM biljaka C 4

EKOFIZIOLOŠKE ODLIKE C-4 i CAM biljaka C 4 biljke sunčanih i toplih staništa, koje se odlikuju karakterističnom anatomijom lista. U asimilacionom tkivu lista nalaze se dvije vrste ćelija: 1. Ćelije mezofila koji su podijeljene na palisadno i sundjerasto tkivo; 2. Ćelije koje spiralno obavijaju sprovodne snopiće tzv. Kranz ćelije.

mnoge C-3 biljke imaju takođe omotač mezofilnih ćelija oko provodnih snopića, ali ove ćelije nemaju, ili imaju malo hloroplasta. Nasuprot tome, u » vijencu « C-4 biljaka nalaze se krupni hloroplasti koji mogu biti postavljeni centripetalno ili centrifugalno ćelije unutrašnjeg omotača, C-4 biljaka imaju zadebljale zidove, prožete suberinom, dok su ćelije mezofila tanke - ćelije » vijenca « i mezofila su povezane dobro razvijenim brojnim plazmodezmama

- hloroplasti u ćelijama sare provodnih snopića imaju posebnu unutrašnju građu -prema strukturi hloroplasta u ćelijama sare C-4 biljke se dijele : - NADP-malatne- kod kojih se nalaze hloroplasti sa pojedinačnim tilakoidima, koji se pružaju dužinom cijelog hloroplasta-nema grana-tzv. agranalni hloroplasti - na pojedinim mjestima se formira grana –samo nekoliko tilakoida bluzu jedan drugom - hloroplasti ćelijske sare se od mezofilnih razlikuje i po enzimima koje sadrže- rubisco i dr. enzimi reduktivno pentoznog ciklusa nalaze se isključivo u hloroplastima ćelija sare (u mezofilnim ćelijama je dominantan enzim karboksilaza fosfoenolpiruvata)

OSOBINA C 3 • Važnije biljne vrste • Prim. prod. fik CO • Rasprostranjenost • Anatomska građa • Fotorespiracija • Optimalna temp. (ºC) 15 - 25 • Transpiracijski koef. C 4 pšenica, krompir 2 kukuruz, ječam kiseline sa 3 C atom velika samo granularni aktivna 450 - 950 kisjeline sa 4 C atoma topla staništa granularni i agranularni nije uočljiva 30 - 47 250 - 350

Metabolizam CO 2 -fiksacija CO 2 iz vazduha i sinteza C-4 organskih kiselina se odvija u mezofilnim ćelijama - reduktivnvi pentozni ciklus se odvija u ćelijama sare - u okviru asimilacije CO 2 razlikuju se sledeće faze: - fiksacija CO 2, karboksilacija fosfoenolpiruvata i sinteza kiselina sa 4 ugljenikova atoma u mezofilu - transport C-4 kiselina u ćelije sare - dekarboksilacija C-4 kiselina, fiksacija CO 2 u reduktivno pentoznom ciklusu u ćelijama sare - transport ostataka sa 3 atoma C u ćelije mezofila - regeneracija akceptora CO 2 u ćelijama mezofila

-procesi u ćelijama mezofila su uglavnom jednaki kod svih C-4 biljaka, a prema C-4 kiselinama koje se transportuju iz mezofila u saru i prema enzimima koji obavljaju dekarboksilaciju C-4 biljke se dijale na 3 grupe: - NADP-malatni tip- sa jabučnom kiselinom i NADP-jabučnim enzimom (kukuruz, šećerna trska…) -NAD tip – sa asparaginskom kiselinom i NAD-jabučnim enzimom (Amarantus edulis, Panicum miliaceum…) - PCK-aspartatni tip- sa asparaginskom kiselinom i fosfoenolpiruvatnom karboksikinazom (PCK) (Panicum maxicum, Spartina …)

Fiiksacija CO 2 i karboksilacija fosfoenolpiruvata - fiksacija CO 2 se obavlja u ćelijama mezofila, koje su zahvaljujući intercelularima, okružene vazduhom - apsorbovani CO 2 se u citoplazmi ćelija rastvara u ugljenu kiselinu (enzim karbodenhidraza) -jon ugljene kiseline se vezuje za supstrat fosfoenolpiruvat –karboksilacija (enzim fosfoenolpiruvat karboksilaza –aktivna ako je fosforilovana, na svjetlosti) - prvi produkt je oksalsirćetna kiselina (oksalo-acetat) koja je toksična i brzo prelazi u dvije kiseline: - kod malatnih biljaka enzim jabučna (malatna) dehidrogenaza čiji je koenzim NADPH redukuje oksaloacetat u jabučnu kiselinu (malat) - kod asparatatnih biljaka aminacijom oksalo-acetata enzimom transaminazom dobija se asparaginska kiselina (aspartat) - karboksilacija PEP u malat kod malatnih biljaka se obavlja u hloroplastu mezofilnih ćelija a NADPH potiče iz svijetle faze fotosinteze -aminacija oksalo-acetata se dešava u citoplazmi mezofilne ćelije - malat odnosno aspartat se zatim transportuju iz mezofilne ćelije u ćelije sare preko plazmodezmi

Biljke sa malatom i NADPH-jabučnim enzimom - malatne biljke imaju samo agranalne hloroplaste u ćelijama sare, u membranama ovih hloroplasta odnos hl a/b je 10, a u mezofilnim ćelijama 2, 5 - malat koji je ušao u hloroplast ćelije sare---pod dejstvom jabučnog enzima, koji ima funkciju dekarboksilaze oksiduje se i gubi CO 2 - --- kao rezultat reakcije dobija se CO 2, NADPH i piruvat (C 3) - CO 2 --- u Kalvinov ciklus za karboksilaciju Ru 1, 5 -bisfosfata - NADPH u Kalvinov ciklus (pošto nemaju grana, PSII je slabo razvijen, samo se obavlja cikličan transport elektrona i proizvodi ATP) - piruvat ide nazad u ćelije mezofila i regeneriše se PEP

Biljke sa aspartatom i NAD-jabučnim enzimom - aspartat koji se ušao u ćelije sare ulazi u mitohondrije i pod dejstvom aspartatne aminotransferaze reaguje sa ketoglutaratom, predaje amino grupu i nastaju oksaloacetat i glutamat - oksaloacetat se redukuje u malat (enzim NADP malatne dehidrogenaze) a zatim se u oksidativnoj dekarboksilaciji pod dejstvom jabučnog enzima vezanog za NAD daje CO 2 i piruvat - CO 2 ulazi u hloroplast--reduktivni ciklus - piruvat u citoplazmi prima –NH 2 grupu prelazi u alanin koji napušta ćeliju sare i ulazi u mezofilnu ćeliju i , ponovo gubi –NH 2 grupu daje piruvat koji se u hloroplastu regeneriše u PEP

Biljke sa aspartatom i PEP-karboksinazom - u mezufilnoj ćeliji se od oksalacetata u citosolu obrazuje aspartat i malat u hloroplastu - slično kao kod biljaka sa aspartatom i NAD-jabučnom kiselinom , aspartat prelazi u ćelije sare, reaguje sa ketoglutaratom (enzim aspartatna aminotransferaza) i dobija se oksaloacetat i glutamat - oksalacetat , pod dejstvom PEPkarboksinaze, za čije je dejstvo potreban ATP, daje PEP i CO 2: - CO 2 ulazi u hloroplast i učestvuje u reduktivnom ciklusu - PEP se vraća u mezofilnu ćeliju - malat iz hloroplasta mezofilne ćelije odlazi u mitohondriju ćelije sare, gdje pod dejstvom NAD-jabučnog enzima daje piruvat, CO 2 i NADH - piruvat kao alanin se vraće iz ćelije sare u mezofilnu ćeliju, ponovo daje piruvat i on se regeneriše u PEP -CO 2 ide u redukcioni ciklus u hloroplastu -NADH se u mitohondriji ponovo oksiduje u elektron-transportnom lancu i doprinosi proizvodnji ATP koji je neophodan za dejstvo PEP-karboksonaze

Regeneracija akceptora CO 2 - proces regeneracije PEP su isti kod sva tri tipa C-4 biljaka i odvija se u mezofilnim ćelijama -enzim piruvat-fosfatna dikinaza PPDK prenosi jednu fosfatnu grupu sa ATP na piruvat , a drugu na jedan neorganski fosfat (Pi) pa nastaju: -PEP, adenozinmonofosfat AMP i pirofosfat PPi - AMP sa još jednim molekulom ATP, pomoću adenilne kinaze proizvodi 2 ADP koji su neophodni kao supstrat u fosforilaciji za sintezu ATP -produkti reduktivnog ciklusa u ćelijama sare se deponuju kao skrob u istim hloroplastima, ili se kao trioze vraćaju u mezofilne ćelije gdje se gradi saharoza. Međutim, saharoza se vraća i ulazi u floem, a skrob se razlaže i takođe transportuje floemom u druge djelove biljke.

Stehiometrija C-4 ciklusa - hloroplasti mezofila: -obavljaju svijetlu fazu fotosinteze i proizvode ATP i NADPH -ne sadrže rubisco enzim, kao ni ostale enzime reduktivnog ciklusa -ATP se koristi isključivo za regeneraciju akceptora CO 2 – PEP, u ciklusu u kome se obnavlja i ADP koji je potreban za fotosintetičku fosforilaciju - NADPH se koristi za reduktivnu sintezu malata kod malatnih biljaka, redukcioni ekvivalent, ugrađen u malat, se troši u hloroplastu sare u reduktivnom ciklusu (kod aspartatnih biljaka prenos redukcionog ekvivalenta nije potreban, jer one imaju granalne hloroplaste i obavljaju necikličan transport elektrona) -u reduktivnom ciklusu u hloroplastima sare troši se na jedan fiksirani CO 2 2 NADPH i 3 ATP kao kod C-3 biljaka, a treba dodati još 2 ATP za regeneraciju PEP

Produktivnost C-4 biljaka Kakav je smisao procesa u kome se CO 2 prvo veže, pa oslobodi, a pri tome se troši znatna energija? -C-4 tip fotosinteze je način kojim se CO 2 koncentriše u hloroplastu, na mjestu gdje se obavlja reduktivni ciklus -to je evolutivna adaptacija biljaka koje žive u toplim i suvim predjelima -glavna posljedica je smanjenje fotorespiracije što dovodi do znatno veće produktivnosti C-4 biljkaka - fotorespiracija kod C-3 biljaka se ne može izbjeći jer fiksaciju obavlja rubisco (kod visokih temperatura fotorespiracija donosi veći gubitak jer je oksigenacija Ru 1, 5 bisfosfat intenzivnija) - C-4 biljke su izbjegle ovaj proces time što fiksaciju CO 2 iz vazduha obavlja PEPkarboksilaza, koja ima visok afinitet prema CO 2 (odnosno prema HCO 3 -) i na koju O 2 ne utiče
 i")
Biljke reaguju na temperaturu Postoje dvije vrste fotosintetske reakcije: • Enzimska (Kalvinov ciklus) i Postoje dvije vrste fotosintetske • reakcije: Elektronska (svjetlosna reakcija) (Kalvinov ciklus) i • Enzimska • Elektronska (svjetlosna Enzimski koraci su veoma reakcija) osjetljivi na temperaturu, dok elektronski Enzimski koraci su su manje veoma osjetljivi, na alitemperaturi, se mogu dok osjetljivi pojavljivati su zbog svojih elektronski manje zavisnosti osjetljivi, aliod seenzimskih mogu koraka. zbog svijih pojavljivati zavisnosti od enzimskih koraka.
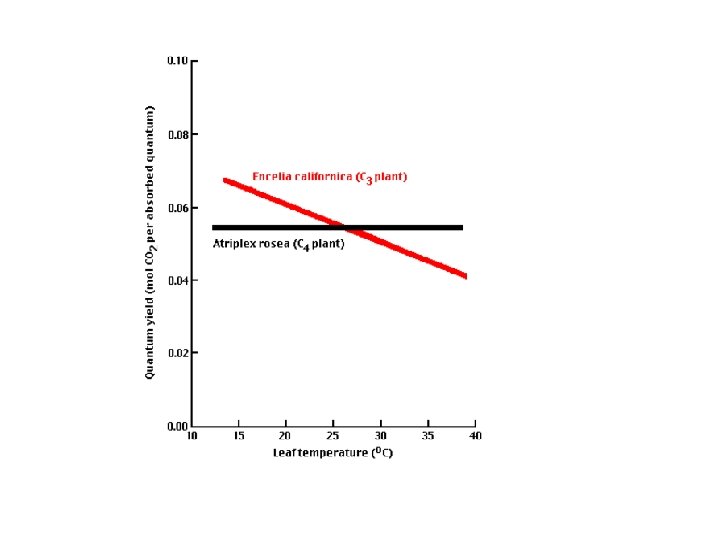

Gdje vode i ugljendioksida ima u izobilju C-3 biljke su dobri konkurenti. Gdje postoji ograničena količina vode i ugljendioksida a da je pri tom temperatura toplija, C-4 biljke su dobri konkurenti.

FIKSACIJA CO 2 KOD SUKULENTNIH BILJAKA -u suvim i vrlo toplim predjelima biljke sa sukulentnim organima čine posebnu ekološku grupu, sa posebnim morfološkim i anatomskim osobinama • KUTIKULA NA POVRSINI- SMANJENJE TRANSPIRACIJE • MESNATA FORMA-AKUMULACIJA VODE • STOME SITNE, MALOBROJNE, UVUCENE I ZATVORENE TOKOM DANA • KORIJEN POVRSINSKI I EKSTENZIVAN • LISTOVI CESTO REDUKOVANI I PRETVORENI U BODLJE • VISOKA AKTIVNOST PEP-KARBOKSILAZE I DEKARBOKSILAZE

• STABLOVE SUKULENTE • LISNATE SUKULENTE

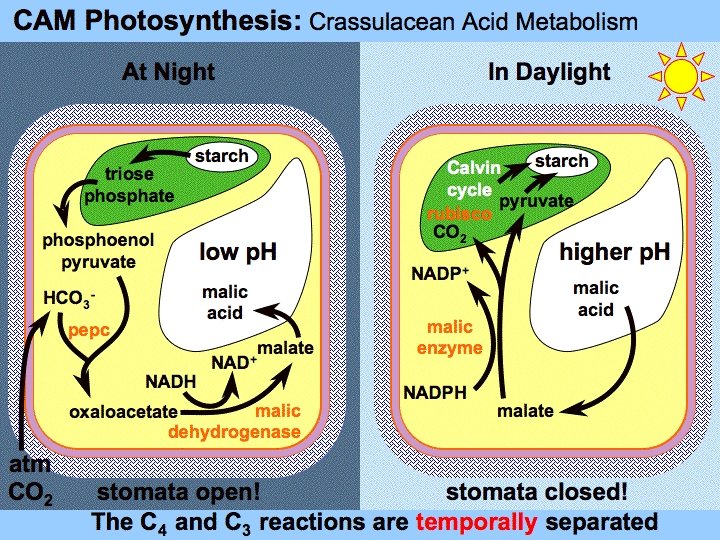
- u fiziološkom pogledu dobro su prilagođene uslovima sredine, jer su razvile sposobnost da ograniče transpiraciju, a da ipak vrše fotosintezu -karateriše ih poseban način koncentrisanja CO 2 u hloroplastima, koji obavljaju reduktivni pentozni ciklus—slično C-4 biljkama -kod C-4 biljaka fiksacija CO 2 iz vazduha i njegova redukcija su odvojeni prostorno u različitim ćelijama -kod sukulenata ova dva procesa su odvojena vremenski---fiksacija CO 2 se obavlja noću, a korišćenje u fotosintezi danju, u istoj ćeliji -sukulentne biljke pripadaju taksonomski različitim grupama (oko 10000 angiospermi, naročito familije Crassulaceae i Cactaceae gdje su svi predstavnici sukulente biljke; familije Liliaceae, nekoliko hiljada orhideja i mnoge agave) - pošto je poseban način fiksacije CO 2 u vezi sa metabolizmom jabučne kiseline, a prvo je proučen kod Crassulaceae, ovaj fiziološki proces je nazvan CAM-metabolizam tj. CAM-biljke (Crassulacean acid metabolism)

Metabolizam CO 2 Fiksacija CO 2 u toku noći - stome su danju zatvorene jer bi gubitak vode mogao biti veći od usvajanja korjenom -zato se stome otvaraju noću, transpiracija je smanjenja ali je i usvajanje CO 2 ograničeno samo na taj period - CO 2 ulazi iz intercelulara u ćelije hlorenhima (asimilaciono tkivo) i u citoplazmi se kao HCO 3 - vezuje za PEP pomoću enzima PEP karboksilaze -nastaje oksalsirćetna kiselina koja se odmah redukuje pomoću NAD-dehidrogenaze i daje jabučnu kiselinu (malat) -kao kod C-4 biljaka samo je malatna dehidrogenaza vezana za citosolni NAD koenzim -PEP vodi porijeklo od skroba, ili drugih ugljenih hidrata, koji se razlažu glikolitičkim putem do PEP - jabučna kiselina tokom noći ulazi u vakuolu (aktivan transport; ATPaza na tonoplastu pumpa protone u vakuolu, a malat ulazi kao posljedica elektrohemijskog gradijenta protona) - danju količina malata u vakuoli iznosi 10 -20 mola g-1, a u toku noći dostiže vrijednost od 200 mola g-1

Metabolizam CO 2 Dekarboksilacija malata u toku dana - akumulacija malata traje tokom cijele noći, ali se brzina, pri kraju tog perioda smanjuje - dok temperatura ne poraste stome su otvorene—za to vrijeme se prima CO 2 iz vazduha i koristi kako za karboksilaciju PEP i proizvodnju malata tako i za karboboksilaciju Ru 1, 5 -BP - postepeno se stome zatvaraju i korišćrnje spoljašnjeg CO 2 se manjuje - počinje dekarboksilacija malata, koji izlazi iz vakuole citoplazmu (enzim NAD-jabučni enzim - malatna dekarboksilaza) - ima mišljenja da se sva tri vida dekarboksilacije malata kod C-4 biljaka javljaju kod raznih CAM biljaka--- u svakom slučaju CO 2 ne može da izađe iz lista, jer su stome zatvorene pa se koristi u hloroplastima za reduktivni ciklus

Regulacija CAM-metabolizma - najvažniji enzim u CAM-metabolizmu je PEP-karboksilaza - vrlo je specifičan kod CAM-biljaka i omogućava regulaciju aktivnosti u ciklusu dan/noć - postoje dva oblika – jedan oblik je danju inhibiran niskom koncentracijom malata - oblik koji je aktivan noću osjetljiv je prema malatu - za jedan CO 2 i CAM-biljke u reduktivnom ciklusu troše 2 NADPH i 3 ATP, treba im 1 -2 ATP za regeneraciju PEP (slično C-4 biljkama) i oko 0, 5 ATP za sintezu polisaharida koji su izvor za PEP tokom noći

UTICAJ INTENZITETA SVJETLOSTI NA PROCES FOTOSINTEZE BILJKA – primarni akceptor sunčevog zračenja, prva karika u lancu ishrane, primarni producent Sunčevo zračenje ima mnogostruko, neposredno i posredno djelovanje na biljke: - fotoenergetski efekat - izvor energije za proces fotosinteze - fotobiološki efekat - stimulus za pokretanje i regulisanje određenih fiziočoških procesa, rast i razviće - fotodestruktivni efekat – preterana količina zračenja izaziva ozbiljna oštećenja kod biljaka - termički efekat – zračenje dugotalasnog dijela spektra
 apsorbovana Sunčeva")
• U fotoenergetskim procesima, fotoreceptorima, odnosno pigmentima lista (hlorofilima i karotenoidima) apsorbovana Sunčeva energija postaje pokretačka snaga mnogobrojnih reakcija i biohemijskih procesa u okviru fortosinteze, na osnovu kojih dolazi do stvaranja visokoenergetskih hemijskih jedinjenja. • Intezitet fotosinteze direktno zavisi od količine apsorbovanog zračenja i efikasnosti glavnih fotosintetičkih receptora.

Fotobiološki efekat se ogleda u djelovanju crvene i plave svjetlosti kao stimulusa za pokretanje i kontrolu različitih bioloških procesa, a prije svega za: - biosinteze enzima, pigmenata, fitohormona; - indukciju fotomorfogenetskih procesa na osnovu kojih se realizuje specifičan oblik i ukupna struktura biljke, što se, u ekološkom smislu, može odrediti i kao formativno dejstvo svjetlosti - usmjeravanje i promjenu pravca rastenja izdanka i listova biljke, odnosno procese fototropizma, naročito heliotropizma, što se ekološki može definisati i kao orjentacijsko dejstvo svjetlosti - regulisanje fotoperiodskih reakcija, kao što su formiranje cvjetova, plodonošenje, opadanje listova, sinhronizovanih sa ritmičkim promjenama kvaliteta i kvantiteta svjetlosti na staništu.

Fotodestruktivno dejstvo sunčevog zračenja - može biti izazvano ili ktratkotalasnim, ultraljubičastim zracima ili iznenadnim izlaganjem biljaka izuzetno intezivnoj bijeloj svjetlosti. Npr. Na šumskim požarištima ili progalama nastalim sječom stabala u šumi, intezivna svjetlost predstavlja prekomjernu fotohemijsku energiju za zeljaste biljke koje su dotle rasle u šumskoj senci. One nisu sposobne da ovu energiju iskoriste, prije svega, u procesu fotosinteze. Uz to se inteziviraju i procesi fotoinhibicije, tako da se smanjuje opšti prinos biomase. Ultraljubičasto zračenje, pak, izaziva fotooksidativne reakcije i fotolezije na biomembranama.

Uticaj svjetlosti na intezitet procesa fotosinteze Fotosintetički proces započinje i odvija se sve dok na staništu ima dovoljno svjetlosti. Pri tome, intezitet fotosinteze je proporcionalan jačini svjetlosti , dok god neki drugi faktor spoljašnje sredine ne ograniči njen dalji rast. Pod uticajem zračenja, asimilacija ugljendioksida najprije raste veoma brzo, dostiže kompenzacijsku tačku i nastavlja da raste, a zatim polagano dostiže maksimalnu vrijednost i tačku zasićenja fotosinteze količinom svjetlosti dostupne na određenom staništu. Odnos između netofotosinteze i zračenja je proporcionalan do tačke zasićenja i predstavljen je krivuljom zasićenja. Sa daljim rastom inteziteta svjetlosti, odnosno u uslovima ekstremnog zračenja, zapaža se veoma slabo ili uopšte nema povećanja fotosintetičkog prinosa.

Određene specifičnosti postoje kada su u pitanju C-3 i C-4 biljke. Za C-3 biljke je karakteristična krivulja rasta fotosinteze hiperboličnog oblika sa izraženim platoom, s obzirom da nema daljeg povećanja fotosinteze poslije postizanja maksimalne vrijednosti, na tački zasićenja. Dalje povećanje količine svjetlosti kod ovih biljaka može dovesti do oštećenja fotosintetičkog aparata i drastičnog smanjenja fotosinteze. Nasuprot tome C-4 biljke se odlikuju visokom kompenzacijskom tačkom i mogućnošću da neprestano, umjereno ili veoma izrazito, povećevaju intezitet fotosinteze, kada se izlože sve jačem svjetlosnom zračenju na njihovim otvorenim staništima. Stoga je svjetlosna krivulja fotosinteze C-4 biljaka manje ili više asimptotskog karaktera, bez upadljivog oblika koji odražava momenat svjetlosnog zasićenja.

Ekofiziološke adaptacije biljaka svjetlosti - Biljke svjetlosti ili heliofite mogu biti jednogodišnje ili višegodišnje zeljaste ili drvenaste biljke koje rastu na otvorenim mjestima (izvan šumskog sklopa), u uslovima pune dnevne osvjetljenosti i maksimalnog Sunčevog zračenja karakterističnog za određenu klimatsku oblast. - Heliofite ulaze u sastav stepa, savana, pustinja, mediteranskih oblasti, stijena, sipara, kao i livada i pašnjaka u nizijskim, brdskim ili visokoplaninskim, prije svega, južno eksponiranim područjima.

Postoji čitav niz morfološko-anatomskih i funkcionalnih prilagođenosti koje omogućavaju biljkama svjetlosti da prežive u uslovima jakog zračenja na otvorenim mjestima. Stablo heliofita, biljaka svjetlosti, je najčešće kratkih internodija i širokog prečnika, nisko, sa obilno razvijenim ksilemom i mehaničkim tkivom i ima bogato razgranatu krunu i lišće usmjereno na sve strane, što se može vidjeti na primjeru vrste Liriodendron tulipifera.

Listovi se obično preklapaju i tako jedan drugog štite, smanjujući dotok Sunčeve energije ili mijenjaju položaj tokom dana, od horizontalnog, ujutru i predveče, do vertikalnog u podne, što im omogućava da smanje količinu svjetlosti kojoj su izloženi. Ukupna površina listova po biljci je redukovana, jer su pojedinačni listovi male površine i izrazito usječeni, za šta kao primjer mogu poslužiti listovi iste vrste, Liriodendron tulipifera.

Ovi sitni listovi su, međutim, debeli, tako da je veći odnos unutrašnje površine mezofila prema spoljašnjoj površini lista. Ova struktuna adaptacija je značajna za visoko produktivnu i efikasnu fotosintezu. Mezofil heliomorfnih listova čini više slojeva kako palisadnog, tako i sunđerastog tkiva.

Korjenovi su brojni i veoma granati, često veoma dugački; povećan je odnos mase korjena prema masi stabla, jer se fotosintezom obrazovana organska supstanca ugrađuje, prije svega, u rast i povećanje korjenovog sistema, na račun slabijeg rasta nadzemnih djelova biljke. Građa korjena Liriodendron tulipifera je i u ovom slučaju ilustrativan primjer.

Jedna od najizrazitijih zaštitinih struktura heliofita je gust dlakavi pokrivač - indumentum koji je često razvijen i na gornjoj i na donjoj strani lista. Gust indumentum značajno mijenja količinu energije koju takav list prihvata, jer je u stanju da reflektuje čak do 70% Sunčevog zračenja vedrog, letnjeg dana. Na taj način, dlakavi listovi pustinjske biljke Encelia farinosa apsorbuju oko 30% punog dnevnog osvjetljenja, dok glatki listovi srodne vrste Encelia californica apsorbuju čak 84% fotosintetički aktivnog zračenja, pri jednakoj koloičini hlorofila po listu navedenih biljaka.

Hloroplasti biljaka svjetlosti su kraći i zaobljeniji, imaju veću zapreminu strome po plastidu, s obzirom na tanke naslage granalnih tilakoida, orjentisanih pretežno u jednoj ravni, a ne u svim pravcima, kao kod hloroplasta biljaka senke. Otuda, oni imaju i relativno malo hlorofila po hloroplastu. Veća zapremina strome po hloroplastu biljaka svjetlosti znači i povećanu količinu proteina, kao i uopšte, metaboličkih enzima. Kako se razvijaju u uslovima intezivne svjetlosti, za heliofite je karakterističan povećan odnos hlorofila a uprema hlorofilu b. Pored toga, kod njih se često javljaju veće količine karotenoida kao zaštitnih pigmenata, što se odražava i na promjenu prevashodno zelene boje lista.
- Slides: 36