EKL BTKLER SSTEMAT BTKLERN SINIFLANDIRMA SSTEMLER BTKLERN SINIFLANDIRMA

ÇİÇEKLİ BİTKİLER SİSTEMATİĞİ BİTKİLERİN SINIFLANDIRMA SİSTEMLERİ

BİTKİLERİN SINIFLANDIRMA SİSTEMLERİ • Sınıflandırma sistemleri tarihi, bilimin tarihi kadar eskidir. Bunun yanında sınıflandırma sadece bilime özgü değildir. Her şey sınıflandırılabilir. Örneğin halkın, çevrelerindeki bitkilere birtakım isimler takmış olmaları bile bir tür sınıflandırmadır. • Günümüze kadar birçok sınıflandırma sistemleri ortaya konulmuş ve sınıflandırmayı yapanların bakış açısına göre belli başlı devrelerde farklı sınıflandırma sistemleri yapılmıştır. • Farklı dönemlerde günümüze kadar sınıflandırma sistemleri 5 grup altında sınıflandırılmıştır.

1 - YAPAY SİSTEMLER M. Ö. 300 -M. S. 1580 • Bu dönemde bitkiler daha çok görünüşlerine göre ve insana sağladığı yararlara göre sınıflandırılmışlardır. • Theophrastus- De Historia Plantaurum adlı eserde 480 bitki çeşidi tanıtılmış, sınıflandırma ağaç, çalı, ayrıçalı, ot, yabani, kültür, yapraklı, yapraksız, tohumlu, tohumsuz gibi karakterler kullanılarak yapılmıştır.


• Dioscorides- Materia Medica eserinde Akdeniz çevresi bitkilerinden 600 bitki tanımlanmıştır.

Albertus Magnus; Bitkileri gövde yapısındaki farklılıklara göre, yapraklı bitkileri Tek çenekliler ve çift çenekliler (otsu/odunsu) olarak ayırmıştır.

2 - Mekanik Sistemler 1580 -1760 • Bu dönemde görünüşün yanı sıra yapısal karakterler dikkate alınmıştır. • Andrea Caesalpina - bitkileri otsu ve odunsu oluş, ovaryumun alt veya üst durumlu oluşu, meyvanın lokulus sayısı gibi yapısal karakterleri almıştır. • Jean Bauhin ayırıcı karakterlerle 5000 bitkiyi tanımlamıştır.

Gaspard Bauhin ilk kez binomial nomenklatürü önermiştir. Tür ile cins arasındaki farkları belitmiştir.

Joannis Raji “Methodus Plantarum” adlı eserinde: • Bitkileri önce otsular ve ağaçsılar olarak ayırmış sonra bunları kendi aralarında tekçift çenek, yaprak, çiçek ve meyve özellikleri v. s. ile tekrar tasnif etmiş ve 18. 000 türün tanımını yapmıştır.

Tournefort sınıflandırmasını otsu ve odunsu, taç yapraklı/yapraksız ve bileşik taç yapraklı olarak yapmış, “Insttitutiones Rei herbariae”adlı eserinde (1700) 698 cins ve 10. 146 türü tanımlayarak ilk kez modern anlamda cins kavramını ortaya atmıştır. • Tournefoth’ 1717 tarihli eserinde tür adı: Gundelia orientalis acanthi aculeati foila capite glabro iken bu isimlendirme sistemine göre adı Gundelia tournefortii

• Bu dönemin en önemli araştırıcısı: Carl Linnaeus’ tur. • Linne sınıflandırmasını üreme organlarına dayandırarak eşeysel bir sınıflandırma anlayışını getirmiştir. Gaspard Bauhinin önerdiği binomial nomenklatürü geliştirerek uygulamıştır. • 1735’de Systema Naturae, 1737’de Genera Plantaurum, 1753’de Species Plantaurum adlı eserlerinde 1000 cins ve 6000 türün tanımını yapmıştır.

3 - Doğal Sistemler 1760 -1880 • Bu dönemde bir veya birkaç karaktere dayalı sınıflandırma yerine olabildiğince çok karakter kullanarak doğal bir sistem oluşturmak amaçlanmıştır. • Michael Adanson (nümerik taksonominin babası) çok sayıda karakteri dikkate alarak bugünkü modern ordo ve familyaları çok yakın gruplara ayırabilmiştir. • De Candolle “Naturalis Regni Vegatabilis” adlı eserde bitkiler vaskular/selüler olarak iki büyük gruba ayrıldıktan sonra Tek/çift çenekliler olarak ayrılmış sonra alt sınıflara bölünmüştür.

• Avusturyalı Stephan Endlicher Kriptogamları fanerogamlardan ayırmıştır. • Bentham-Hooker Genera plantaurum adlı eserlerinde bitkiler önce Dicotyledonae/polypetalae, Dicotyledone/gamopetalae gibi büyük gruplara ayrılmış, daha sonra çiçek özelliklerine göre seri-takım ve familyalara bölünmüştür. Sistem tesadüfen Ranales ile başlar. Sistemleri Filogenetik sistemlerle çok yakın ilgili olması dolayısıyla ilgi görmüştür. • Bu dönemin son araştırıcısı W. Hoffmaeister, sisteminde Thallophyta, Bryophyta, Pteridophyta, Gymnospermae ve Angiospermae gibi büyük bölümlerin ayrılmasında embriyolojik karakterleri kullanmıştır.

4 - Filogenetik Sistemler 1800 -günümüz Bu dönem Darvin’in “Türlerin kökeni” adlı eserini yayınlamasıyla başlamıştır.

• Türler arasındaki akrabalık derecelerine veya daha genel anlamda soy ağacına dayanan sınıflandırmalar yapılmış, doğada var olan akrabalık ilişkilerin bulunması hedeflenmiştir. • Geçek akrabalığa dayanan benzerliklerle, uyum sonrası benzerliklerin birbirinden ayrılması gerektiğini, kullanılan benzerliklerin gerçek bir soy ortaklığına dayanan benzerlikler olması gereğini vurgulamıştır. • Darvin’in bu görüşleri “Evrim Kuramı” döneminin doğmasına neden olmuştur.

• Darvin Siteminde bazı kriterleri almıştır. • Türler sabit olmayıp zamanla değişebilir. Zaman ve mekan içinde değişen formlardır. • Türler fertler topluluğu değil, Populasyonlar topluluğu olarak kabul edilir ve türün esas ünitesi fert değil populasyondur. • Türleri ayırımda fizyolojik durumun (üreme gibi) dikkate alınması gerekir

İlk filogenetik sistemi ortaya koyan araştırıcı August W. Eichler dir. Buna göre: Cryptogamae Phanerogamae Gymnospermae Thallophyta Bryophyta Angiospermae Monocotyleae Dicotyleae Choripetalae Sympetalae Pteridophyta

• Eichleri takiben ortaya atılan sistem Engler-Prantl sistemidir. Alman Ekolü: • Angiosperm’lerde Dikotiller ilkel kabul edilen Amentiferae ile başlar. Monokotiller ise Pandanales’le başlar. Nedeni: Pandanales çiçek durumu tipik bir Gymnosperm kozalağına benzer. Amntiferae de ise tozlaşma anemogam, çiçekler tek eşeyli ve periant indirgenmesi vardır. Bununla beraber evrim sırası: Periantsızlar Sepaloid brakteliler Sepalleri ve petalleri serbest olanlar Sepalleri bulunan ve petalleri birleşik olanlar Buna göre sistem: Apetalae Ranales Tubuliflorae şeklinde sıralanır.

• Diğer önemli filogenetik sistem Bessey’dir • Bu sisteme göre: Angiospermae; Bennetitales takımından monofiletik olarak türevlenmiştir. Bu Nedeni Bennettitales’in çiçek durumunun manolyadaki çiçek durumuna benzer olmasıdır. • Bessey kaktüs ağacı şeklinde bir sistem düşünür ve bunun altına Ranales’i koyar ve burada çok parçalı, serbest ve homojen yapıları ilkel kabul eder.

Bessey sınıflandırmasını yaparken birtakım prensipleri dikkate almıştır: A. Genel Prensipler 1. Evrim hem pozitif hem negatif yöndedir. 2. Homojen yapılar ilkel, heterojen yapılar gelişmiştir v. s. B. Bitkilerin Genel yapısı ile ilgili prensipler 1. Kolleteral diziliş dairesel dizilişten ilkeldir. 2. Ağsı damarlanma, paralel olandan ilkeldir. 3. Ağaç durumu otsu durumdam ilkeldir. C. Çiçeğin yapısı ile ilgili prensipler 1. Aktinormf simetri zigomorfdan ilkeldir. 2. Stamenlerin ayrı oluşu ilkelliktir vs.

• Engler-Prantl ve Bessey sistemleri birer ekol oluşturmuşlardır. Bu nedenle: • Amentifera’yı ilkel kabul eden Engler-Prantıl sistemine “ENGLERİAN TİP”, • Ranales’i ilkel kabul eden Bessey sistemine de “RANALİAN TİP” sistemler denir.
 sistemidir. • Temel olarak Bentham-Bessey ekolüne aittir.")
Diğer önemli filogenetik sistem Hutchinson (1926 -1960) sistemidir. • Temel olarak Bentham-Bessey ekolüne aittir. • Angiospermlerin monofiletik olduğunu, atalarının Gymnosperm’lerin bir grubu olduğunu kabul eder. Dikotilleri başlıca odunsular (Lignosae) ve otsu olanlar (Herbaceae) olarak ikiye ayırır. • Herbaceae Ranales ile, • Lignosae Magnoliales ile başlar

Hutchinson da sistemini kurarken bazı prensipleri kabul etmiştir. Örneğin: 1. Ağaç ve çalılar otsulardan ilkeldir. 2. Suda yaşayanlar, karadakilerden ilkeldir. 3. Sarmal diziliş dairesel dizilişden ilkeldir 4. Basit yapraklar bileşiklerden, hermafrodit çiçekler tek eşeylilerden ilkeldir 5. Aktinorf çiçekler zigomorf dan ilkeldir 6. Hipogin durum perigin ve epiginden ilkeldir 7. Bileşik meyvalılar tek meyvalılardan ilkeldir vs.

5 - Çağdaş Sistemler • Bir türün yayılış alanının farklı yerlerindeki populasyonlar incelendiğinde, bunlar arasında birtakım farkların olduğu görülür. Bu da “Populasyon sistematiği” diye bir akımın doğmasına neden olmuştur. • Türlerin tanımlanması çağdaş sistematikte, populasyonların tanımlanması şeklinde kabul edilmektedir. Çünkü bireylerin zaman ve mekan içinde sınırlarının bulunmasına karşılık, populasyonlar süreklidir. • Populasyon sistematiği klasik taksonominin bir seçeneği değil, onun daha da geliştirilmiş şeklidir. • Klasik taksonomi pratik olarak canlıları tanımlamayı amaçladığından ve morfolojik özellikler göre yapılabildiğinden uzun süre geçerliliğini koruyacaktır.
 Sayısal taksonomi organizma grupları arasındaki benzerliğin sayısal değerlendirmesi ve onları")
Nümerik Taksonomi (sayısal Taksonomi) Sayısal taksonomi organizma grupları arasındaki benzerliğin sayısal değerlendirmesi ve onları bu benzerliklere dayanarak gittikçe yükselen takson katları içersinde sıralamak olarak tanımlanabilir. Bunun için önce benzerliklerin ölçülmesi ve sonra da sınıflama hiyerarşisinin yapılması gerekir. Sayısal taksonomi tamamen fenetik yani taksonlara ait gözlenmiş ve kayıt edilmiş karakterlerin gösterdiği benzerliklere dayanır. Filogenetik olasılıklara dayanmaz. Sayısal taksonomi yeni verileri ortaya koymaz ve üstelik yeni bir sınıflama sistemi de değildir. Ancak verileri yeni bir organizasyonla düzenleyen bir metottur ve bu metot sayesinde bir sınıflama elde edilir.

Sınıflandırma Kuramları 1. Esasçılık: Aristo mantığına dayanan ve Linne tarafından kabul edilen görüştür. Türlerin değişmezliğini kabul eder. 2. İsimcilik: doğada sadece bireylerin bulunduğunu, diğer tüm sistematik kategorilerin insanlar tarafından oluşturulduğuna inanır. Akrabalık kavramına değil sadece türler arasındaki benzerliklere dayanır. 3. Amprikcilik: Taksomomist için, ayrıca bir sınıflandırma teorisine gerek olmadığına, yeterli miktardaki karakterler alındığında bu sistemin kendiliğinden ortaya çıkacağına inanır. (Nümerik Taksonomi) 4. Filogenetik ilişkilere dayanır. Soy ağaçları yapar. 5. Gelişmiş sınıflandırma: Doğada var olan grupların benzerliklerini evrim ile açıklayarak bulmaya çalışan görüştür.
’e göre yüksek bitki ve hayvanlarda evrim başlıca 4")
EVRİM ve TÜRLERİN FARKLILAŞMASI Stebbins (1959)’e göre yüksek bitki ve hayvanlarda evrim başlıca 4 olayın sonucudur: 1. Mutasyon 2. Genetik Rekombinasyon / Varyasyon 3. Doğal seleksiyon 4. İzolasyon
 biyolojik tür oluşumunun esasını,")
Türlerin oluşumunda üreme izolasyonu en kritik aşamadır. • Dobzansky (1950) biyolojik tür oluşumunun esasını, genetik olarak ayrılabilen Mendel populasyonları arasındaki üreme izolasyonunun başarıya ulaşmasına bağlamaktadır. • Genel olarak 2 tip tür oluşumu vardır. Ani tür oluşumu ve dereceli tür oluşumu. Bununla beraber her iki tür oluşumu şekli de ancak üreme izolasyonu ile başarıya ulaşır.


Ani tür oluşumu İyice belirgin coğrafik ve ekolojik farklılıkları olan ve sadece gen değişimi sınırlandırılmış çiftler arasındaki kromozom sayısı bakımından farklılaşan gruplardır. Kromozom sayısı ve yapısındaki ani bir değişim sonucu ortaya çıkarlar. Bu ani değişimse genellikle poliploidinin sonucudur ve bu poliploidler eğer başarılı olurlarsa belli ekocoğrafik bölgeleri işgal ederler. Dolayısıyla bu oluşum simpatriktir denilebilir.

Dereceli tür oluşumu • Aynı kromozom sayısına sahip; iyi belirlenmiş morfolojik, ekolojik ve coğrafik farklılıkları olan ve doğal ya da doğal olmayan şartlarda sadece sınırlandırılmış gen değişimleri ile bağlı gruplardır. • Dereceli tür oluşumu mutasyon, rekombinasyon, seleksiyon ve izolasyon sonucu oluşan farklılıkların birikimi sonucu, adım ilerleyen bir oluşumdur. Bunda da coğrafik izolasyonun etkinliği tartışmasız çok önemlidir. Bu nedenle bu oluşum allopatriktir denilebilir. • Tüm dereceli tür oluşumlarının simpatrik olarak başladığı sonraları allopatrik izolasyonla ayrıştığı gözlenmektedir.

Poliploidlerin sistematikteki önemi: Poliploidlerin, diploid soylarının ortadan kalkmasına neden olan uygunsuz şartlarda bile yayılabilmelerini sağlayan daha geniş toleransları ve dayanıklıkları vardır. Ancak bilindiği gibi poliploidlerin diploidlere oranla kromozom sayısı bakımından tekrar değişmeleri mümkün olmadığından, koşullardaki yeni ve önemli bir değişime uyum göstermeyip ortadan kalkabilirler. Bu nedenle de poliploidler evrimsel açıdan ölü son olarak isimlendirilirler.

Poliplopidlerin oluşumu ile ilgili çok sayıda kuram vardır: • Genel olarak ani sıcaklık değişimlerinin polyploidiyi doğurduğu, • ekvatordan kutuplara doğru gidildikçe poliploidinin arttığı yolundaki görüşler ağırlıkta ise de bazı ayrıcalıkları da vardır. • Poliploidinin evrendeki işlevi gerçekten çok önemlidir.
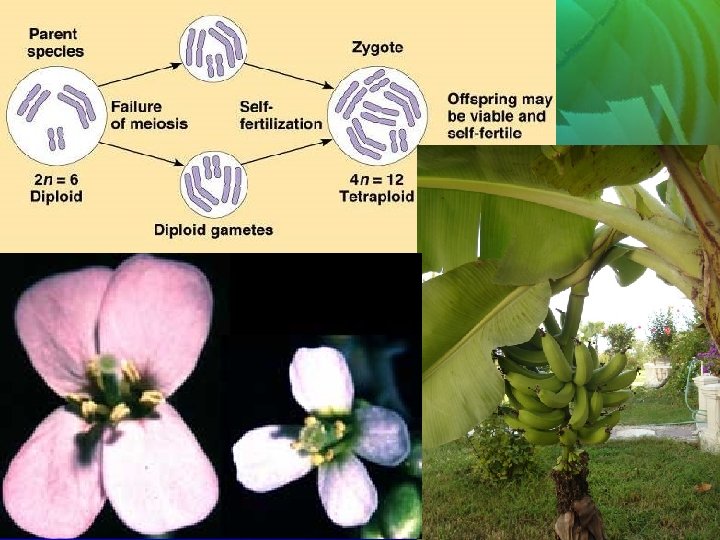

Poliploidinin istenmeyen yönleri: • Poliploidler ileri derecede kısırlılık gösterir. • Diploidlere oranla çok daha az değişkendir. • Poliplidlerden çekinik bir mutasyon ortaya çıkma şansı çok azdır. • Çoğunluk eşeysiz ürediklerinden (apomiksis), yeni genetik kombinasyonlar oluşturamazlar. • Diğer taraftan hayvanlarda ise hemen hiç poliploidiye rastlanmaz.

Yeni türlerin Oluşumunda 3 aşama ! Yeni Türlerin Oluşumu Seleksiyon İzolasyon Varyasyon • Varyasyonlar genetik kökenlidir ve yeni karakterlerin oluşumunda rol oynar. • Oluşan karakterlerin devamlı olup olmayacağına seleksiyon karar verir. • Eğer karakter adaptif bir karakterse ve yaşam savaşında ait olduğu bireye üstünlük sağlıyorsa kalıcı olur ve devam eder.

• Gerçekte değişim populasyon düzeyinde ise anlamlıdır. • Değişime uğrayan bir populasyonun evrim açısından başarılı olması ise üreme izolasyonuna bağlıdır. • Bu izolasyon ani tür oluşumundaki gibi birdenbire olabildiği gibi, dereceli tür oluşumundaki gibi araya birtakım engellerin girmesi ve böylece diğer populasyonlardan soyutlama şeklinde, uzun süreli de olabilir.

İzolasyonda amaç: Populasyonlar arasındaki gen alışverişini önlemek ve böylece; Sadece kendi içinde gen değişimlerinde bulunan populasyonların zamanla farklılaşmasını sağlamaktır.

Taksonomik açıdan önemli bazı İzolasyon mekanizmaları 1. Coğrafik ve Ekolojik İzolasyon 2. Mevsimsel ve Zamansal izolasyon 3. Mekanik ve Etolojik izolasyon 4. İç Engeller

1 - Coğrafik ve Ekolojik İzolasyon • İzolasyon mekanizmaları arasında etkinliği en fazla olanıdır. • Çeşitli bölgeler değişik iklim ve ekolojik şartlar altında bulunduğundan saf coğrafik ve saf ekolojik izolasyon arasında kesin bir sınır çizmek olanaksızdır. Bununla beraber; • Allopatrik populasyonlarda izolasyon hem coğrafik hem ekolojik olabildiği halde, • Simpatrik populasyonlarda izolasyonu sağlayan etmenlerden birisi de ekolojik izolasyondur. Simpatrik türlerin çoğu niteliklerini ekolojik izolasyon sonucunda kazanırlar.

• Populasyonlar coğrafi olarak izole edilirken önce ekolojik ve morfolojik farklılaşma oluşur. Yani ilkin ekotipler ortaya çıkar ve bu ekotiplerin ikinci bir farklılaşması ile de coğrafik ırklar (subspesiesler) oluşur. • Bu olay, yeni türlerin, bir türün yayılış alanının merkezinden çok, daha aşırı şartlara sahip kenar kısımlarında oluşma eğiliminde olmasıyla uyum içersindedir. Eğer bu kenar populasyonlar bölgesel olarak izole olurlarsa, aynı türe ait diğer populasyonlardan ekolojik olarak farklılaşmış olacaklardır.

• Populasyonlar eğer çok geniş bir yayılım alanına sahiplerse, coğrafik izolasyon pek etkin olmaz. Ancak geniş bir alanda yayılan bir türün çeşitli populasyon serileri bulunabilir. Bu nedenle de aşırı uçtaki üyeler etkin bir şekilde izole olabilirler. • Gen merkezi kuramına göre de yeni türlerin, bir türün yayılış alanının kenarında oluştuğu yani, tür farklılaşması merkezden ziyade kenar bölgelerde olmaktadır. Bunda Ana merkezdeki genler nispeten belirli olmasına karşın, kenarlara doru gidildikçe temas edilen populasyonlarla olabilecek olası bir gen alışverişi sonucu populasyona değişik genlerin girmesi rol oynar.

Coğrafik ve Ekolojik izolasyona örnekler • Dağ floralarındaki mikroendemiklerin durumu. Populasyonlar daima bölgesel ve ve az-çok ayrılmış birimlere bölünmüştür. • Etkin bir izolasyon oluşturan ada floraları; Havai adaları, Madagaskar adaları, hatta Avustralya kıtasının uzun süreden beri ana karadan ayrı kalmış olmaları, endemikçe zengin olmaları bu savı doğrulamaktadır.

• Ülkemizde en fazla endemik türün Irano. Turanian bölgede bulunduğu, bunu Akdeniz bölgesinin izlediği ve en az endemik türün ise Euro-Sibirian bölge olduğu bilinir. • Tersiyerin alt- mioseninde ülkemizin homejen bir iklime ve floraya sahip olduğu bilinmektedir. • Homogen olan bu flora günümüz Karadeniz bitkilerine benzer mezofil bitkilerden oluşmaktadır. Buna göre: • İklimin, günümüz Karadeniz iklimine yakın bir iklim olduğu söylenmektedir.

Ülkemizde Endemizm • Ülkemizin Tersiyerin Plioseninde geçirmiş olduğu tektonik değişimleri takiben, topoğrafik ve iklimsel değişmeler doğal floralarda da büyük değişiklikler neden olmuştur. • Anadoluda son 100 bin yıllık devre içinde vejetasyon önemli ölçüde değişmiştir. • Karadeniz bölgesindeki flora aynı kaldığı halde diğer bölgelerde büyük değişiklikler oluşmuştur. • Özellikle Iran-turan bölgesindeki endemik tür fazlalığında bölgenin dağlık olmasının da rolü büyüktür. • Karadeniz bölgesinde ise dağların paralel uzanması yeterince izolasyonu sağlamamış görünmektedir.

Eurosibirian Bölge Iran Turan Bölgesi Akdeniz Bölgesi

Anadolu Diyagoneli

Anadolu Diyagoneli

2 - Mevsimsel ve Zamansal İzolasyon Mevsimsel izolasyon daima ekolojik ve coğrafik izolasyonu kuvvetlendirici yönde etki eder. Bitkilerin çiçeklenme zamanlarındaki farklılıklar sonucu da izolasyon gerçekleşmektedir.

Örneğin: G. doğu Amerika'da yol kenarlarında yayılış gösteren; Lactuca canadensis Lactuca graminifolia (yazın çiçek açar) (ilkbaharda çiçek açar) türleri aynı kromozom sayısına sahip ve interfertil oldukları halde farklı mevsimlerde çiçek açtıklarından aralarında mevsimsel izolasyon vardır.

Kaliforniya’da yayılış gösteren Pinus radiata P. muricata hem ekolojik tercihi hem de çiçeklenme zamanlarındaki farklılıklar sonucu izole olmuşlardır.

• Ülkemiz Crocus’ larında da tayin anahtarlarında çiçeklenme zamanındaki farklılıklar kullanılmıştır. • Allopatrik olan Galanthus nivalis ssp. reginae-olgae sonbaharda, Galanthus nivalis ssp. cilicicus kış ortasında, Galanthus nivalis ssp. nivalis ise ilkbaharda çiçek açar.

Ülkemizde bulunan, birbirleriyle interfertil olan ve aynı kromozom sayısına sahip; Cyclamen cilicicum Sonbaharda çiçek açar. Cyclamen coum İlkbaharda

Zamansal izolasyon ise polenlerin günün çeşitli saatlerinde yayılmasına ya da stigmanın günün değişik zamanlarında polen kabul etmesine dayanır. Çok fazla etkin bir izolasyon mekanizması değildir. Örneğin: Agrostis stolonifera polenlerini sabah 10. 30 -11. 30 arasında yayarken, Agrostis tenuis ise öğleden sonra 13. 00 -14. 30 arasında yaymaktadır.

3 - Mekanik ve Etolojik İzolasyon • Mekanik izolasyon çiçeklerin yapısal karakterlerindeki farklar üzerine kurulmuştur. Orchidaceae ve Asclepiadaceae familyalarında olduğu gibi bu tip izolasyon bitkilerde oldukça etkendir. • Etolojik izolasyonda ise iki tür arasında çapraz tozlaşma olasıdır, ancak tozlayıcının sadece bir türe özgü olması nedeniyle oluşamaz. Bu durumda etolojik ve mekanik izolasyon iç içe girmiş olup hangi durumda hangisinin daha etken olduğunu söylemek güçtür.

Örneğin: • Anthirrinum majus ve A. glutinosum arasındaki ve kokudaki farklılıklar nedeniyle çiçek seçiciliği sağlanır. • Ophrys’ lerde herbir türü özel bir arı türünü çektiğinden, bunlarda türler arası çaprazlama çok güç olmaktadır. Ophyris lutea Ophyris sicula

Çiçeklerle böcekler arasındaki karşılıklı ilişki oldukça ilginçtir. • Bazı cinsler, türlerin adaptasyondaki başarısını sadece farklı böceklerle değil, aynı zamanda farklı hayvan gruplarıyla tozlaşmalarına borçludur. Örneğin; Aquilegia’ nın arılarla, kelebeklerle, bülbüllerle ve böceklerle tozlaşan türleri vardır.

Mekanik ve etolojik izolasyon belki populasyonların ayırımında rol oynamayabilir, ancak türlerin evriminde başarılı rolleri olmuştur denilebilir.

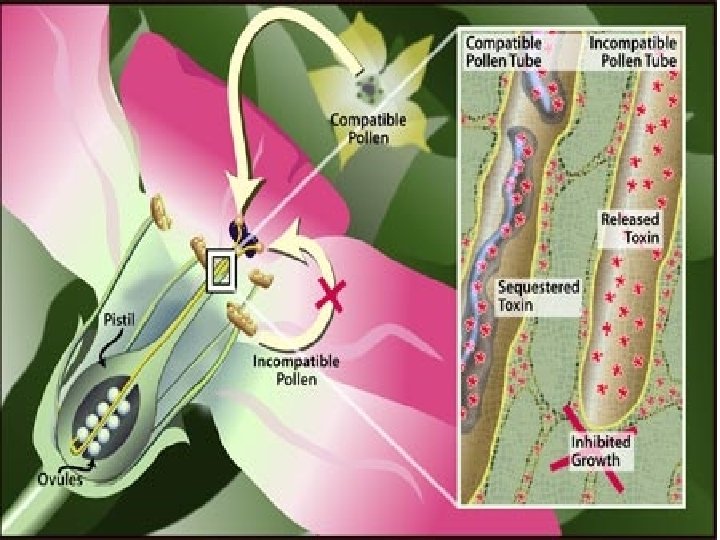
4 - İç Engeller • Melez oluşumunu önleyen ya da oluşanların verimliliğini azaltan doku farklılıklarının meydan getirdiği içsel engellerdir. Polenlerin diğer bir türün stigması üzerinde çimlenememesi; çimlenirse, polen tüpünün ovaryuma erişememesi; şayet erişip döllerse embriyoyu besleyecek endospermin oluşmaması bir iç engeldir. • Nicotiana (tütün) bitkisinde self sterilite (kendine kısırlık) uzun bir allel serisi tarafından sağlanmaktadır.
- Slides: 61