Ecology Physiology of Fungi Fungal Nutrition LEC6 Fungal

















 plant parasites give out slender lateral")

 usually consists of two parts, a constricted region which")

- Slides: 22
Ecology & Physiology of Fungi Fungal Nutrition LEC=6
Fungal Nutrition Yeasts and fungi have relatively simple nutritional needs and most species would be able to survive quite well in aerobic conditions if supplied with glucose, ammonium salts, inorganic ions and a few growth factors. Macronutrients, supplied at millimolar concentrations, comprise sources of carbon, nitrogen, oxygen, sulfur, phosphorus, potassium and magnesium; and Micronutrients, supplied at micromolar concentrations, comprising trace elements like calcium, copper, iron, manganese and zinc, would be required for fungal cell growth
Some fungi are oligotrophic, apparently growing with very limited nutrient supply, surviving by scavenging minute quantities of volatile organic compounds from the atmosphere.
Being chemoorganotrophs, fungi need fixed forms of organic compounds for their carbon and energy supply. Sugars are widely utilized for fungal growth and can range from simple hexoses, like glucose, to polysaccharides, like starch and cellulose. Some fungi can occasionally utilize aromatic hydrocarbons (e. g. lignin by the white-rot fungi). Fungi are non-diazotrophic (cannot fix nitrogen) and need to be supplied with nitrogenous compounds, either in inorganic form, such as ammonium salts, or in organic form, such as amino acids. Ammonium sulfate is a commonly used nitrogen source in fungal growth media, since it also provides a source of utilizable sulfur.
Phosphorus is essential for biosynthesis of fungal nucleic acids, phospholipids, ATP and glycophosphates. Hence, the phosphate content of fungi is considerable (e. g. in yeast cells it accounts for around 3– 5% of dry weight; the major part of this is in the form of orthophosphate (H 2 PO 4−), which acts as a substrate and enzyme effectors). The fungal vacuole can serve as a storage site for phosphate in the form of complexed inorganic polyphosphates (also referred to as volutin granules). Both nitrogen and phosphorus availability may be growth limiting in nature.
Filamentous fungi have evolved a number of biochemical and morphological strategies allowing capture of often poorly available phosphorus within the natural environment. Plants exploit such efficiency during symbioses between their roots and certain mycorrhizal fungi. The major storage form of phosphorus in plants is phytic acid (myo-inositol hexadihydrogenphosphate), which is poorly utilized by monogastrics (e. g. humans, pigs, poultry), and fungal (and yeast) phytases have applications in reducing phytate content of foods and feeds.
Concerning requirements for minerals, potassium, magnesium and several trace elements are necessary for fungal growth. Potassium and magnesium are macroelements required in millimolar concentrations, primarily as enzyme cofactors, whereas other microelements (trace elements) are generally required in the micromolar range. These include Mn, Ca Fe, Zn, Cu, Ni, Co and Mo. Toxic minerals (e. g. Ag, As, Ba, Cs, Cd, Hg, Li, Pb) adversely affect fungal growth generally at concentrations greater than 100 μM.
Fungal growth factors are organic compounds occasionally needed in very low concentrations for specific enzymatic or structural roles, but not as energy sources. These include vitamins (e. g. thiamine, biotin), purines, pyrimidines, nucleosides, nucleotides, amino acids, fatty acids and sterols. For fungi to have a growth factor requirement, this indicates that cells cannot synthesize the particular factor, resulting in the curtailment of growth without its provision in culture media. Some fungi (e. g. Aspergillus niger, Penicillium chrysogenum) have very simple nutritional needs and are able to synthesize their own growth factors from glucose.
Element Carbon Common sources Sugars Cellular functions Structural element of fungal cells in combination with hydrogen, oxygen and nitrogen. Energy source Hydrogen Protons from acidic Transmembrane proton motive force vital forfungal nutrition. Intracellular environments acidic p. H (around 5– 6) necessary for fungal metabolism Oxygen Air, O 2 Substrate for respiratory and other mixed-function oxidative enzymes. Essential for ergosterol and unsaturated fatty acid synthesis Nitrogen NH 4 + salts, urea, Structurally and functionally as organic amino nitrogen in proteins and amino acids enzymes Phosphorus Phosphates Energy transduction, nucleic acid and membrane structure Potassium K+ salts Ionic balance, enzyme activity Magnesium Mg 2+ salts Enzyme activity, cell and organelle structure Sulfur Sulfates, methionine Sulfydryl amino acids and vitamins Calcium Ca 2+ salts Possible second messenger in signal transduction Copper Cupric salts Redox pigments Iron Ferric salts. Fe 3+ is Haem-proteins, cytochromes chelated by siderophores and released as Fe 2+ within the cell Manganese Mn 2+ salts Enzyme activity Nickel Ni 2+ salts Enzyme activity Molybdenum Na 2 Mo. O 4 Nitrate metabolism, vitamin B 12
Mode of Nutrition: Absorption The mode of nutrition or the matter in which fungi "eat" is called absorption. Among eukaryotes, absorption is unique to the fungi. Fungi obtain their food by transporting it through their cell walls. In order to eat, the spores that give rise to fungi must be dispersed to a location where there is food and after the spore germinates, the mycelium of the fungus must grow into its food. Another word, usually fungi must live in their food if they are to eat.
If the food is composed of simple molecules such as glucose or sucrose, soluble food can be immediately transported through their cell walls. However, most food that a fungus might consume is composed of complex, organic compounds, e. g. , cellulose, lignin, pectin, starch, etc. , which is insoluble. In order for this food to be utilized by the fungus, it must be broken down into simpler molecules that can be transported through their cell walls.
Unlike green plants, which use carbon dioxide and light as sources of carbon and energy, respectively, fungi meet these two requirements by assimilating preformed matter; carbohydrates are the preferred nutrient source. Fungi can readily absorb and metabolize a variety of soluble carbohydrates, such as glucose, xylose, sucrose, and fructose, but are also characteristically well equipped to use insoluble carbohydrates like starches, cellulose, hemicelluloses, and lignin. To do so, they must first digest these polymers extracellularly. Saprobic fungi obtain their food from dead organic material; parasitic fungi do so by feeding on living organisms (usually plants), thus causing disease.
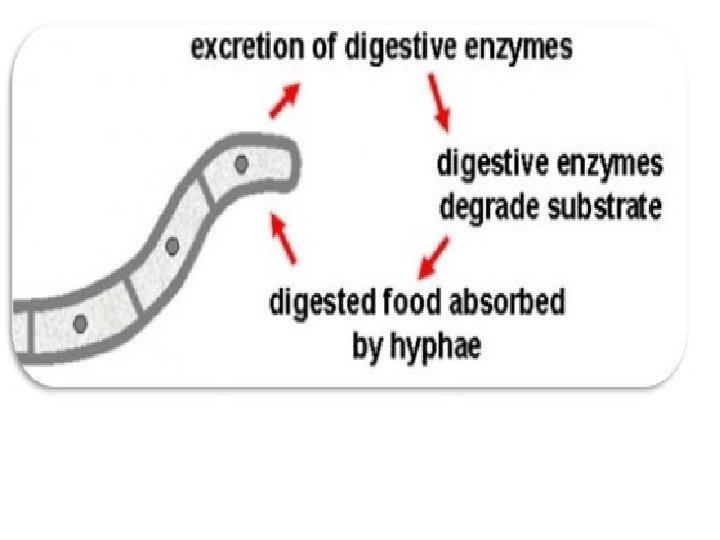
The fungus breaks down the complex material by secreting digestive enzymes through their cell wall that will digest the complex organic compounds and convert them into simple molecules that can readily be transported through their cell walls. For example, If a fungus is growing in wood, digestive enzymes would be secreted from the fungus, into the wood, and break down the complex compounds of wood, e. g. cellulose and lignin into simpler materials, such as simple sugars, which then can be transported into the mycelium
Illustration of the process of absorption, the mechanism by which fungi consume their food.
Food must enter the hyphae in solution, and, since most fungi have no special absorbing organs, the entire mycelia surface is capable of taking in materials dissolved in water. Some fungi, however, produce special root like hyphae, called rhizoids, which anchor the thallus to the growth surface and probably also absorb food. Many parasitic fungi are even more specialized in this respect, producing special absorptive organs called haustoria in Fig. 1.
Although this process may seem very different than our own means of obtaining food. It is not that different. The essential difference between fungi and animal digestive systems is that fungi digest their food first and then "eat" it, while animals eat their food before digesting it. It is important to understand here that different kinds of fungi will secrete only a specific number of different enzymes. This means that they can only "eat" certain materials. A summary of absorption is illustrated in Fig.
Mechanism of Nutrition The whole mycelium may have the power to absorb these nutrients or this task may be assigned to special portions of the mycelium. In saprophytic fungi the hyphae (Mucor mucedo) or rhizodial hyphae (Rhizopus stolonifer) come in intimate contact with nutrients in the substratum (A) and absorb soluble smaller molecules such sugars and amino acids. Insoluble complex substances such as proteins, lipids and Poly are first broken into soluble monomeres (digested) by secreting extra-cellular enzymes and then absorbed. The fungal hyphae secrete enzymes which convert insoluble complex food materials in the substratum to soluble ones. The latter are then absorbed by direct diffusion either through the hyphal walls of the hyphae that penetrate the substratum or by the rhizoidal hyphae. The mycelium of the parasites is rarely ectophytic but frequently it grows inside the host. The hyphae either ramify in the intercellular space between the host cells (D) or penetrate into the host cells (G). The former are called intercellular hyphae and the latter intracellular hyphae. The intercellular hyphae obtain nutrition through the cell walls or membranes of the host cells. This they do by secreting an enzyme upon the plasma membrane of the host cell. It makes the membrane more permeable to the contained solutes. The latter diffuse out and are absorbed by the hyphal walls. The hyphal walls of the intracellular hyphae come in direct contact with the host protoplasm (G) and obtain food by direct diffusion.
The intercellular hyphae of some highly specialised (obligate) plant parasites give out slender lateral outgrowths. The hyphal outgrowth punctures the host cell wall making a minute pore through which it enters the host cell. Within the host cell, it enlarges to form a globose (D), lobed (B), or branched (F) absorptive organ.
This type of feeding organ of the parasitic fungi is called a haustorium. It is markedly specialised in structure to absorb nutrition from the host tissues. The haustonum is intracellular and thus robs the host of its food without killing it. Haustoria are characteristic of obligate parasites. They vary in shape and size in different fungi. In Albugo the haustorium is a button-like (D) or spherical structure. Peronospora parasitica has sac-like haustoria (E) in the leaf cells of Capsella. Peronospora calotheca produces branched filamentous haustorium in the stem cells of Galium (F). Erysiphe graminis forms an elongated branched haustorium inside the host cell (B).
Each haustorium (Fig. 1. 16) usually consists of two parts, a constricted region which is in the form of a narrow penetration tube and the expanded or branched region on the host cell. The penetration tube is usually clasped by a ‘collar’ of host wall material. The enlarged or expanded region of the haustorium causes Invagination of the cytoplasmic membrane of the host cell. The latter remains closely appressed to the wall of the haustorium. There is a zone of apposition enclosing the haustorium between the fungal wall and the unbroken cytoplasmic membrane of the host. Its origin is in dispute. The secretion from the haustorium upon the plasma membrane of the host makes it permeable to solutes contained in the sap cavity. They diffuse out and are then absorbed by the haustorium parastic fungi do not produce haustoria in artificial cultures. The haustona are also not produced by fungi which live as parasites on animals.
The fungi, as mentioned above, are unable to synthesize sugars from carbon dioxide and water. They, however, can synthesize from soluble sugars the more complex carbohydrates which I the chief components of their cell walls. They are also able to synthesize proteins and eventually protoplasm if supplied with carbohydrates and simple nitrogen compounds such as ammonium salts. Besides ammonium salts, they can absorb and utilize many complex but soluble organic nitrogenous compounds.