Doal Baklk Sistemi Ouzcan ZEREN Alaaddin KILI Muhammed

Doğal Bağışıklık Sistemi Oğuzcan ÖZEREN Alaaddin KILIÇ Muhammed RAHMAN Mustafa Alpagut KAPLAN İrem BOĞA
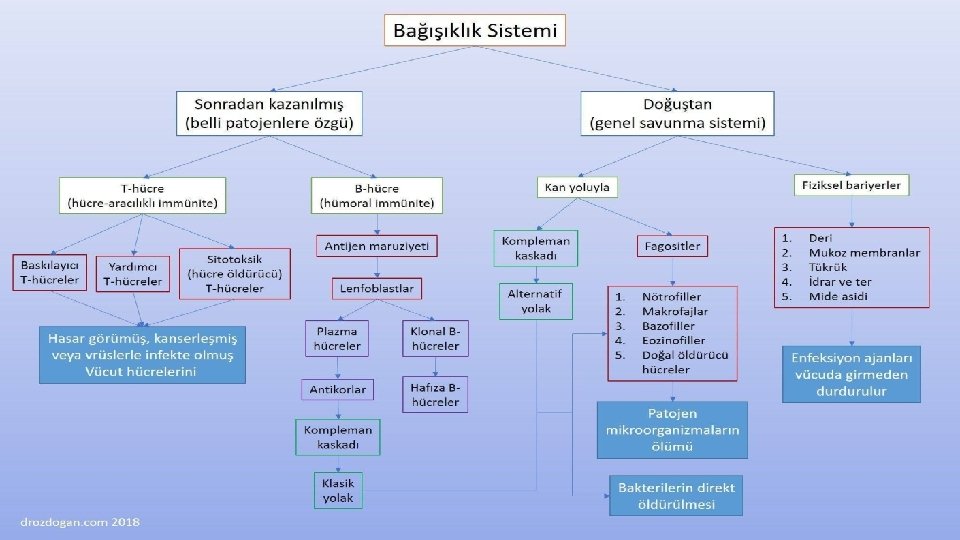
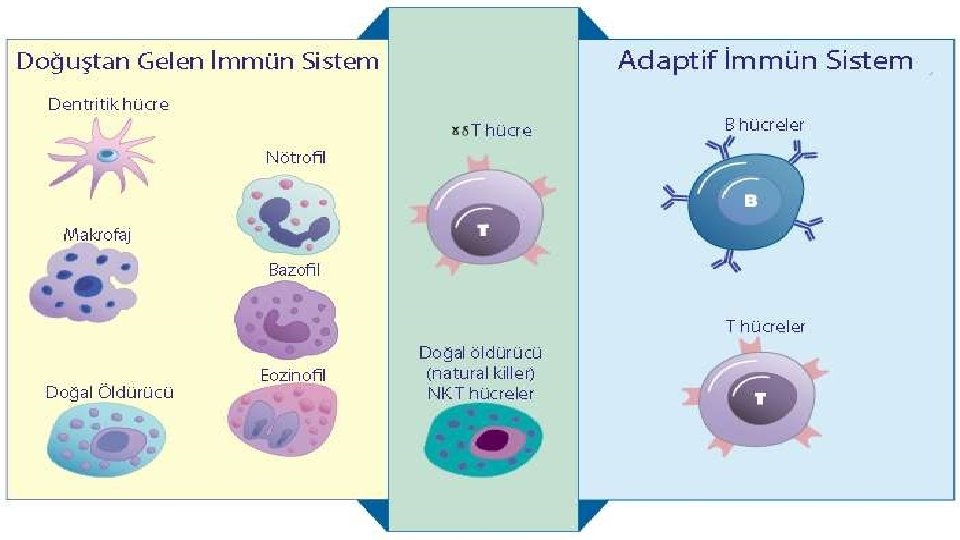

Doğal bağışıklık uyarılma gereksinimi gösteren edinsel bağışıklıktan farklı çalışır. Konak hücrede bulunmayan fakat farklı mikroorganizmalarda ortak olan yapıları tanır. Ancak doğal bağışıklık edinselden farklı olarak hafıza özelliği göstermez. (metillenmemiş Cp. G gibi) Fakat bu iki sistem aynı ana karta bağlı iki farklı bilgisayar gibidir.

Doğal bağışıklık enfeksiyöz etkenler için yaşamsal öneme sahip ve enfeksiyözitelerini sağlayan bölgelerini tanır. Bu sayede bir mutasyon dahi gelişse etkenin doğal bağışıklıktan kaçabilmesini sağlamaz.
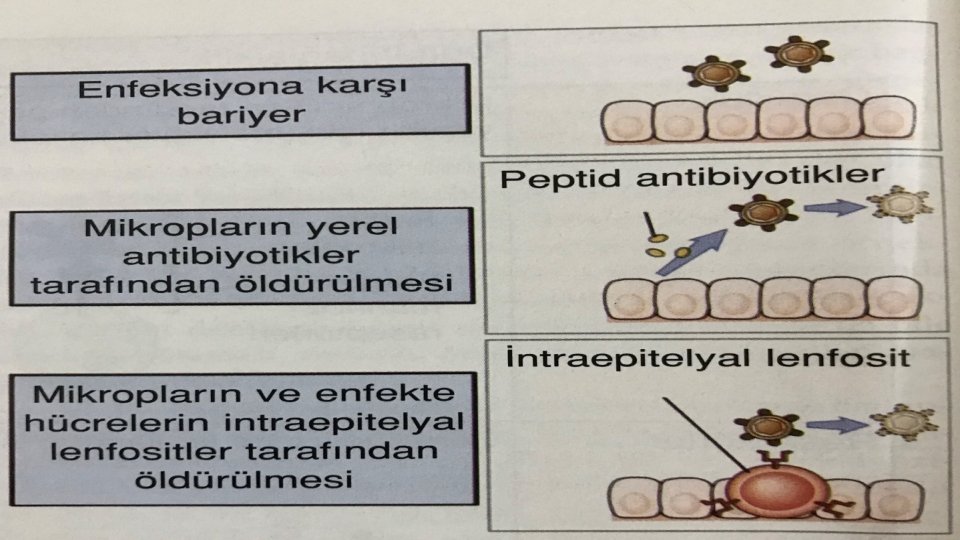

Epitel Tabakası Bir etken için ilk engel, dış ortamla temas halinde olan deri, solunum ve sindirim sisteminin hepsi için geçerli olan, epitel tabakasıdır. Epitel tabakası bariyer görevi görür. Peptit yapılı antibiyotikler üretir. Aynı türden organizmalarda ortak yapıları tanıyan intraepitelyal lenfosit içerir.

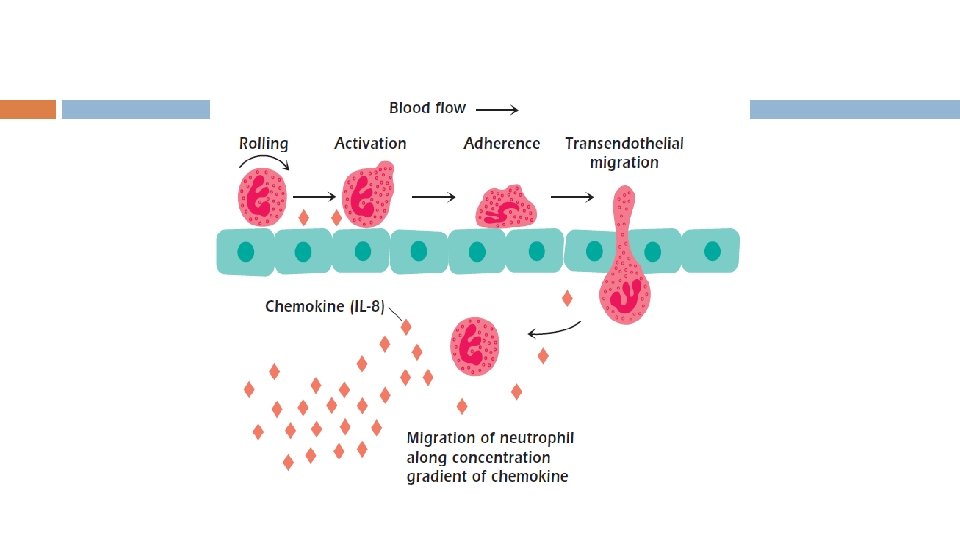
Fagositik Hücreler Nötrofiller sitokinlerce uyarılıp damar dışında da enfeksiyon odağına gidebilen, mantarlarda dahil birçok etkene karşı etkili, kanda en yoğun bulunan hücrelerdir. Monositler de kenoatraktanların etkisiyle damar dışına çıkabilir ve daha sonra dokuda yerleşik kalabilir. Bu durumda makrofaj adını alır.

Fagositik Hücreler Nötrofil ve makrofaj dokudaki enfeksiyonu Toll Benzeri Reseptör denilen farklı bölgelerce tanır. Bu bölgeler aracılığıyla nükleer faktör Kb üretilir ve transkripsiyon faktörü aktive edilerek sitokinler ve antimikrobiyal proteinlerin üretimi sağlanır.

Fagositik Hücreler Makrofajlar interferon gama gibi sitokinler için reseptör taşır. İNF gama fagositik hücrelerin mikrobisidal etkinliklerini güçlü biçimde uyarma özelliği taşır.

Fagositik Hücreler Etkeni tanıyan nötrofil ve makrofaj en son aşamada fagositoz işlemini gerçekleştirir. Fagosit oksidaz, nitrik oksit sentaz, lizozomal proteaz gibi bir takım enzim mikrobiyal proteini parçalar. Fakat çok güçlü yanıt oluşan olaylarda bu enzimlerin ürünleri konağa zarar verebilir. Kalıtsal olarak ortaya çıkan fagosit oksidaz enzim eksikliği kronik granülomatoz hastalık olarak bilinen immün yetersizlik hastalığına neden olur.

FAGOSIT AKTIVASYONU VE EFEKTOR FONKSIYONLARI

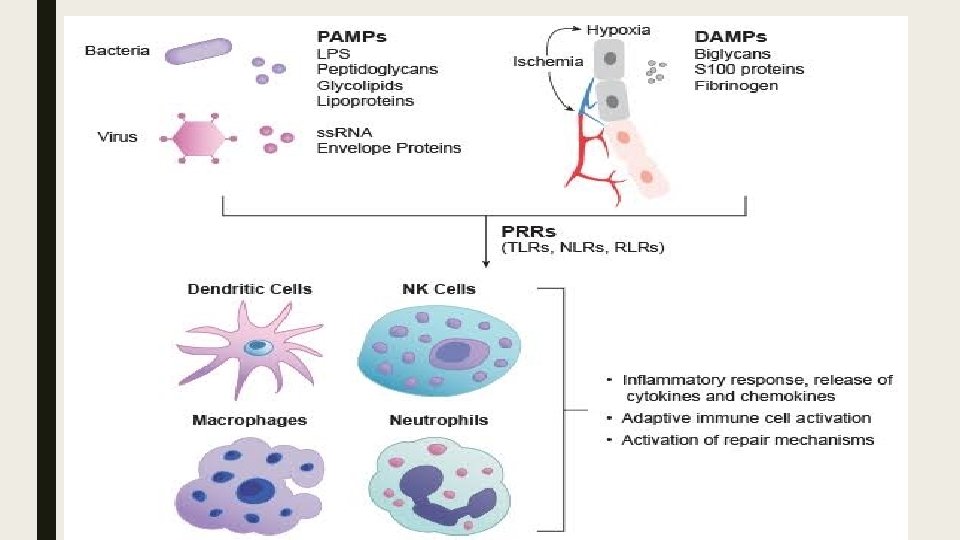
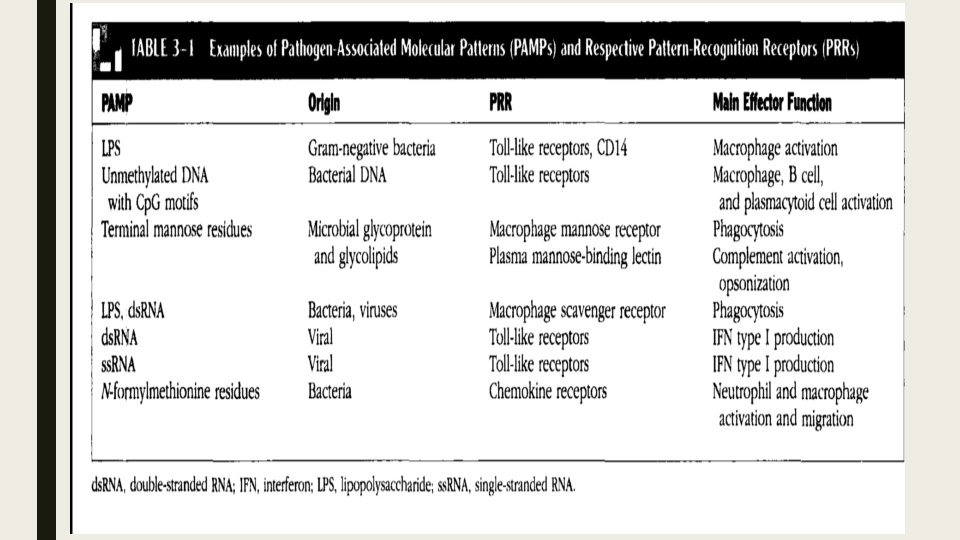
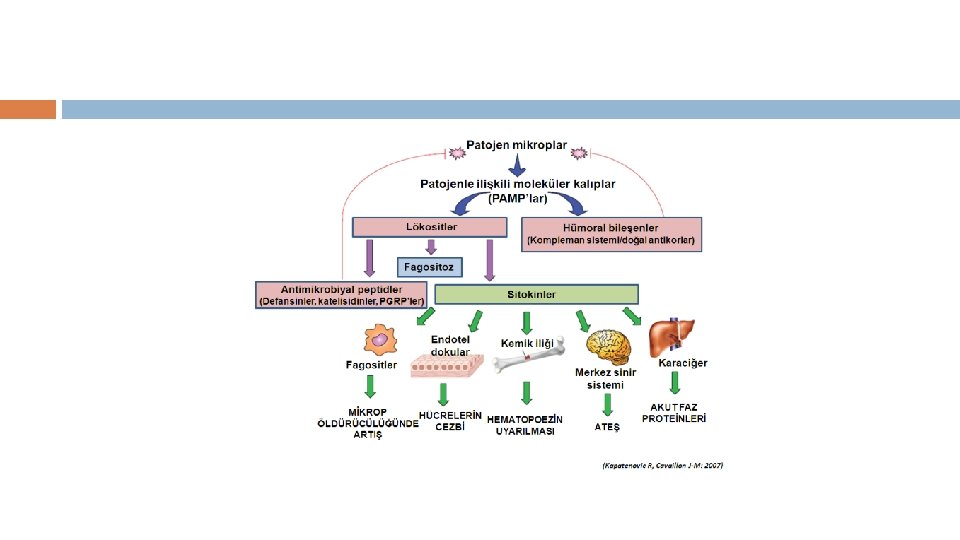
■ Notrofil ve Makrofajlar bir kac yolla aktive edilebilir. Bunlardan birincisi patojenlerin yuzeylerinde bulunan ozgun yapilari taniyan reseptorler yoluyla aktivasyondur. ■ Bu reseptorlerin yuzeyinde bagisiklik yanitinda oncu ve son efektor gorevi goren molekuller var. ■ Bagisiklik hucrelerinde bulunan reseptorlerin tanidigi yapilar istilaci mikroplarin yasamasi icin gereklidir ve patojenle ilgili molekuler paternler (PAMPs) diye tanimlanirlar.
 diye adlandirilir. ■ PAMP’lar degisik")
■ Dogal bagisikligin PAMP’lari taniyan reseptorleri PRRs (Patern-taniyan reseptorler) diye adlandirilir. ■ PAMP’lar degisik mikroorganizm cesitleri arasinda belirgin degisiklik gosterir ama ayni mikroorganizma sinifi arasinda degisiklik gostermezler. Bu karakteristikleri, kisitli sayidaki germ-line kodlu PRR’larin degisik mikrobiyal infeksiyonlari tespit etmelerine izin verir. ■ PAMP’lar mikrobiyal yasam icin olmazsa olmaz oldugu icin, PAMP’lardaki mutasyon veya delesyonlar olumculdur. Bu da mikroplarin PAMP mutasyonlarina ugrayarak dogustan gelen immun sistemden kacma olasiligini onemli derecede
 Dendritik Hucrelerin yuzeyinde ' doku nobetcileri'")
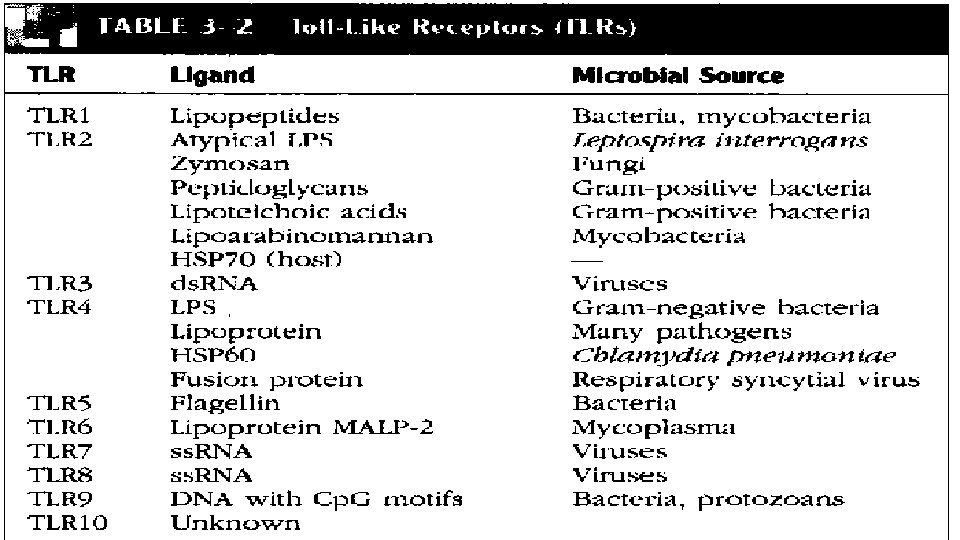
■ PRR'larin bir cok sinifi makrofaj ve (DCs) Dendritik Hucrelerin yuzeyinde ' doku nobetcileri' olarak bulunur, olasi bir mikrobiyal patojen invazyonuna karsi periferik dokulari surekli monitorize ederler. ■ Toll-benzeri reseptorler (TLR-s) PRR'larin buyuk bir sinifidir ve Ekstraselluler Losinden zengin tekrar domaini (LRR) ve de intraselluler Toll/Interlokin-1 (IL-1) reseptor domaini ile karakterize olurlar. ■ Çok farkli ve degisik PAMP'lara yanit vermekte rol alan en az 10 degisik TLR sinifi vardir. ■ Bazi durumlarda, PAMP'larin taninmasi ve fagosit aktivasyonu icin ardindan gelen intraselluler sinyaller aksesuar proteinlerin varligini gerektirir.

■ Mikroplarin fagositlere PRR araciligiyla baglanmasi istilaci mikroorganismanin fagositoz olayini baslatir ve ardindan fagolizozom ile bunun oldurulmesi gelir. ■ PRR araciligiyla fagositlerin aktive edilmesi bircok efektor molekulu de indukler, induklenebilen Nitrik Oksit gibi (NOS) ve direkt olarak mikrobiyal patojenleri oldurebilen diger antimikrobiyal peptidler gibi. ■ Ayni zamanda, mikrobiyal proteinlerden elde edilen peptitler sonradan kazanilmis bagisiklik sistemini uyarmak icin T-hucrelerine sunulur. ■ Bundan baska, PRR’lar tarafindan sinyal ileti yollarinin aktivasyonu, hastanin istilaci mikroplara karsi, hem dogal (inflamatuar sitokinler, buyume faktorleri, kemokinler, proteolitik enzimler) hem de sonradan kazanilan bagisiklik (Major histokompatibilite kompleksleri, ortak uyarici molekuller) yanitlarini duzenleyen genlerin induklenmesine neden olur.

■ Makrofaj aktivasyonunun 2. yontemi Th-1 lenfositlerin IFN-y uretimi yoludur. ■ Makrofaj ve Notrofiller Immun komplex ve komplament fragmanlari araciligi ile de aktive edilebilirler. Bu yol notrofillerin primer aktivasyon yoludur ve bu notrofiller inflamasyon bolgesine ilk sirkule olan lokositleri temsil eder.

Apoptotik hücrelerin fagositozu sürecinde doku makrofajları alternatif aktivasyona uğrar. apoptotik hücrelerin B 2 glikoprotein 1 ve vitronektin reseptörleri gibi yüzey reseptörlerine bağlanmasıyla uyarılır. Makrofajların alternatif aktivasyonu, TGF B, IL-1 O ve IL-1 reseptör antagonisti dahil olmak üzere bir dizi anti-inflamatuar molekülün üretimine yol açar ve akut enflamasyonu durdurmak ve konağı doku hasarından korumak için önemlidir.

• IL-10, pro-enflamatuar sitokinlerin üretimini inhibe eder, reaktif oksijen ara ürünlerinin salımını bastırır ve majör histo-uyumluluk kompleksi (MHC) sınıf II ve kostimülatör moleküllerin ekspresyonunu down regulasyonunu düzenler. • IL-IRa, makrofajlarda bulunan IL-1 reseptörüne rekabetçi bir şekilde bağlanır, böylece IL-1'in proinflamatuar etkisini inhibe eder. • TGF- başka bir pro-enflamatuar sitokinin, tümör nekroz faktörünün (TNF) üretimini inhibe eder ve matris proteinlerinin makrofajlar tarafından ekspresyonunu teşvik ederek hücre dışı matris birikimine, yara iyileşmesine ve fibroza katkıda bulunur

Dentritik hücreler

• Plazmasitoid DCler, belirli virüslere ve diğer mikrobiyal uyarıcılara (bunlara plazmasitoid interferon üreten hücreler de denir) yanıt olarak büyük miktarlarda tip I IFN (alfa beta) salgılama konusunda benzersiz kapasiteye sahiptir. • DC'ler, saf T hücrelerini aktive edebilen en güçlü APC'ler olmasına rağmen, artık DC'lerin de. T hücresi anerjisinin indüksiyonu, T hücresi apoptozu, T düzenleyici hücreler ve T hücreleri tarafından immünodüzenleyici sitokinlerin üretimini içeren çeşitli mekanizmalar yoluyla tolerojenik olabileceği açıktır
 içeren çok sayıda sitoplazmik")
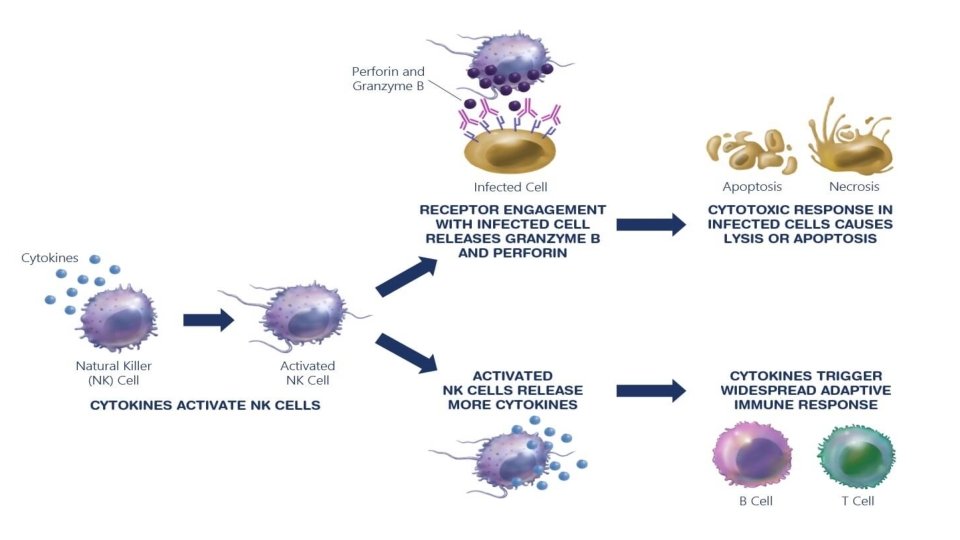
Doğal öldürücü hücreler • NK hücreleri, proteolitik proteinler (perforin, granzimler) içeren çok sayıda sitoplazmik granülün varlığı ile karakterize edilen büyük lenfositlerdir. Antijene özgü reseptörlerden yoksundurlar, ancak bazı tümör hücreleri ve virüsle enfekte hücreler gibi anormal hücreleri öldürebilirler. " • NK hücrelerinin aktivasyonu, aktive edici ve inhibe edici hücre yüzeyi reseptörleri aracılığıyla düzenlenir. İnhibitör reseptörler, çoğu hücre tipinde eksprese edilen kendi sınıf I MHC moleküllerine bağlanır.

Fibroblastlar • • Fibroblastlar, enflamatuar yanıtın efektör kolunda aktif bir rol oynar. Bağ dokusu, daha düşük transformasyon kapasitesine sahip olgun fibroblastlar ve birkaç farklı hücre soyuna farklılaşabilen olgunlaşmamış fibroblastlar (mezenkimal fibroblastlar) dahil olmak üzere farklı fibroblast soylarının bir karışımını içerir. Çok potansiyelli bir karaktere sahip fibroblast öncülleri de kanda dolaşır; kemik iliğinin stromal hücreleri ile benzerliklerinden dolayı mezenkimal kök hücreler olarak adlandırılırlar. • İnflamasyon sırasında, doku makrofajları tarafından üretilen proinflamatuar sitokinler, kemokinler, prostaglandinler (PGE 2) ve proteolitik enzimler üretmek için olgun doku fibroblastlarını aktive eder. Aktif doku fibroblastlarının kapatılamaması, kemokin ve proinflamatuar sitokinlerin sürekli aşırı ekspresyonu ve bunun sonucunda lökositlerin sürekli olarak toplanması yoluyla kronik enflamasyona yol açan bir mekanizma olarak önerilmiştir.

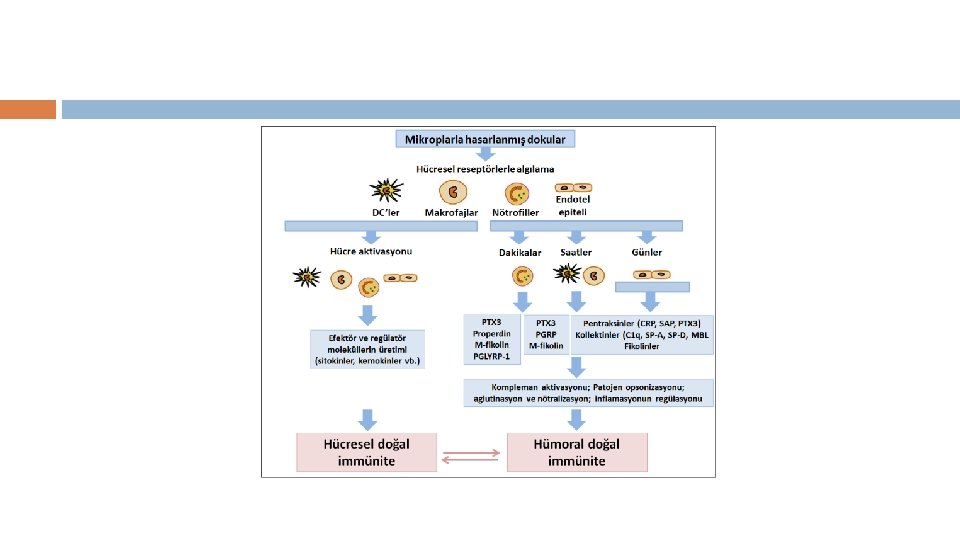
DOĞAL BAĞIŞIKLIĞIN MOLEKÜLLERİ KOMPLEMAN SİSTEMİ

• Kompleman sistemi: aktivasyondan sonra fagositozun desteklenmesi, hücre lizisi ve inflamasyonun uyarılması dahil olmak üzere önemli efektör fonksiyonlarına aracılık eden bir dizi ürün oluşturmak için etkileşime giren birkaç normal inaktif plazma proteininden oluşur. • Kompleman aktivasyonu, pıhtılaşma sistemine benzer bir enzimatik kaskad oluşturan sıralı proteolitik adımları içerir.

• Kompleman aktivasyonunun üç ana yolu vardır. • Alternatif yol, kompleman proteinlerinden biri olan C 3 b'nin hücrelere doğrudan bağlanmasını içerir. • Klasik yol, C 1’in immünoglobulin G’nin CH 2 alanlarına veya bağlı antijene sahip Ig. M'nin CH 3 bölgelerine bağlandığı daha karmaşık bir aktivasyon modunu içerir. • Klasik yolda yer alan aynı proteinler, mikrobiyal glikoproteinler ve glikolipidler üzerindeki mannoz kalıntılarına bağlanan bir plazma proteini (mannoz bağlayıcı lektin) tarafından antikorların yokluğunda aktive edilebilir; bu lektin yolu olarak bilinir.
 Lektin patojen yüzeyindeki mannoza bağlanır Patojen yüzeyler")
Antijen: antikor kompleksler (patojen yüzeyler) Lektin patojen yüzeyindeki mannoza bağlanır Patojen yüzeyler

• Kompleman aktivasyonunun üç yolu, iki fragmana bölünen merkezi bir komplement bileşeni olan C 3'te birleşir. Daha büyük parça (C 3 b) kovalent olarak hücrelere bağlanır, burada fagositozu uyarmak için opsonin görevi görür ve sonraki C 5 b nesli ile C 5'i aktive eder. C 5 b, hücre zarları (membran atak kompleksi, MAC) üzerinde toplanan tamamlayıcı proteinler C 6, C 7, C 8 ve C 9 kompleksinin oluşumunu başlatır ve hedef hücrenin parçalanmasına neden olan bir gözenek oluşturur.

İnflamasyonun peptid mediatörleri Fagositlerin çağrılması Fagositlerdeki kompleman reseptörlerine bağlanır Patojenlerin opsonizasyonu İmmunkompleksler in uzaklaştırılması MEMBRAN ATAK KOMPLEKSİ Belli patojenlerin ve hücrelerin öldürülmesi


• Kompleman aktivasyonu sırasında, daha küçük kompleman fragmanları (C 3 a, C 4 a, C 5 a) dolaşıma salınır. Anafilotoksinler olarak da bilinirler, mast hücrelerinin ve nötrofillerin aktivasyonu ve artmış vasküler geçirgenlik dahil olmak üzere çeşitli proinflamatuar etkiler uygularlar. • Komplemanın bir başka işlevi, antijen antikor komplekslerine bağlanmak, böylece bunların fagositler tarafından çözünmesini ve temizlenmesini teşvik etmektir. Son zamanlarda, komplemanların fagositik sistem tarafından apoptotik kalıntıların temizlenmesinde rol oynadığı gösterilmiştir.

• Kompleman biyolojik aktivitelerine komplement fragmanlarının membran reseptörlerine bağlanması aracılık eder. C 3 fragmanları için reseptörler en iyi karakterize edilir. Tip 1 kompleman reseptörü (CR 1, CD 35) hemen tüm kan hücreleri tarafından eksprese edilir ve C 3 b kaplı hücrelerin fagositozunu destekler. Eritrositler üzerinde ifade edilen CRI, bağlı C 3 b ile dolaşımdaki bağışıklık komplekslerine bağlanır. Bu şekilde, dolaşımdaki eritrositler, bağışıklık komplekslerini, eritrosit yüzeyinden çıkarıldıkları ve temizlendikleri karaciğere ve dalağa taşıyabilirler.
, lenf düğümü germinal merkezlerinin")
• Tip 2 kompleman reseptörü (CR 2, CD 21), lenf düğümü germinal merkezlerinin B lenfositlerinde ve foliküler DC'lerinde bulunur. Ana işlevi, antijen tarafından B hücresi aktivasyonu için koreseptör olarak hareket etmek ve antijen-antikor komplekslerinin germinal merkezlerde yakalanmasını uyarmaktır. Tip 3 ve tip 4 kompleman reseptörleri, integrin ailesinin üyeleridir ve doğuştan gelen bağışıklık hücreleri tarafından ifade edilir (nötrofiller, NK hücreleri, mononükleer fagositler), CR 3 veya CR 4'ün bağlanması, bu hücrelerin aktivasyonunu ve opsonize edilmiş hücrelerin fagositozunu destekler.

• Kompleman eksiklikleri bir dizi patolojik durumla ilişkilidir. Klasik yol bileşenlerinin (C 1 q, C 1 r, C 2 ve C 4) genetik eksiklikleri, SLE benzeri bir tabloya neden olabilir. Bu, apoptotik hücrelerin ve dolaşımdaki immün komplekslerin temizlenmesinde erken tamamlayıcı bileşenlerin rolü ile ilgili olabilir. • C 3 eksikliği, ciddi piyojenik enfeksiyonlarla ilişkilidir. • Terminal kompleman bileşenlerin (C 5 - C 9) defektleri, Neisseria meningitidis dahil olmak üzere, yaygın Neisseria enfeksiyonları riskinde artış ile ilişkilidir.

• Kompleman kaskadının aktivasyonu, normal konakçı hücreler üzerinde aktivasyonu engelleyen mikrobiyal hücreler ve antijen antikor kompleksleri üzerindeki kompleman aktivasyonunun süresini sınırlayan hücre membran proteinleri tarafından düzenlenir. C 1 inhibitörü (C 1 INH), klasik yolun başlatıcısı olan C 1'in proteolitik aktivitesini düzenler. Bu proteinin eksikliği, otozomal dominant kalıtsal anjiyonörotik ödem e neden olur.

• Bir dizi membran proteini (membran kofaktör proteini, MCPj tip 1 kompleman reseptörü, CRl; bozunma hızlandırma faktörü, DAF) ve plazma proteini, faktör H, normal memeli hücrelerinin yüzeylerinde birikmesi durumunda C 3 b'nin aktivasyonunu engeller. • DAF'ın hücre zarı ile bağlanması için gerekli bir enzim eksikliği, eritrositlerin yüzeyinde düzensiz kompleman aktivasyonu nedeniyle tekrarlayan intravasküler hemoliz atağı ile karakterize edilen paroksismal noktürnal hemoglobinüri adı verilen bir hastalığa neden olur. Benzer şekilde, nadir görülen faktör H eksikliği, C 3 tüketimine ve glomerülonefritlere yol açan aşırı bir alternatif yol aktivasyonu ile karakterizedir.

• Son deneysel gözlemler, TGF-p ve IL 10, 25, 26 gibi anti enflamatuar sitokinleri salgılayan düzenleyici T hücrelerinin indüksiyonunda komplemanların bir rolü olduğunu göstermiştir. Bu veriler, kompleman sisteminin bağışıklığın düzenlenmesinde daha geniş bir rol oynadığını göstermektedir.

DOĞAL BAĞIŞIKLIK

MANNOZ BAĞLAYICI LEKTİN Mannoz bağlayıcı lektin kollajen benzeri bir serum proteinidir. Akut faz yanıtı proteinidir Kollektinler ailesindendir Karaciğer tarafından sentezlenir. Karbonhidrat tanıma parçasına sahiptir mannoza seçici olarak bağlanır. Komplemanın lektin yolunu c 1 q üzerinden aktive eder. Mikroorganizmaların yüzeyindeki şeker gruplarına bağlanarak onların makrofajlar tarafından fagositozunu sağlar. Yani opsonizasyon ve kompleman aktivasyonu aracılığıyla doğal immuniteye katkıda bulunur.
 ve SAP(serum amiloid peptid) Crp ve SAP pentraksinler ailesinin üyesi")
CRP (c reaktif protein) ve SAP(serum amiloid peptid) Crp ve SAP pentraksinler ailesinin üyesi olan plazma proteinleridir. İnflamasyonun akut fazında karaciğer tarafından bolca üretilirler. indüksüyonu IL-6 tarafından transkripsiyonel düzeyde regüle edilir, bu etki IL-1β ile artırılır. Mikrobiyal membranda bolca mevcut olan fosforilkoline bağlanırlar. Klasik kompleman yolunu aktive edebilirler. Nötrofil için opsonin görevi görebilirler

KISA PENTRAKSİNLER UZUN PENTRAKSİNLER SAP: Serum amiloid P CRP: C reaktif protein PTX: Pentraksin NP: Nöronal pentraksin NPR: Nöronal pentraksin reseptörü PENTRAKSİN AİLESİ

DEFENSİNLER Antimikrobiyal peptid ailesinin üyesidir. Mukozada saptanan , bakteri hücre zarını delerek etkili olan bir peptiddir. Deri ve mukoza epiteli gibi primer engeller üzerinde salgılanarak konak dokularının patojenlerle kolonizasyonunu önlerler. Granülositlerin antimikrobiyal etkilerine katkıda bulunurlar.

SİTOKİN ve KEMOKİNLER Doğal ve edinsel bağışıklıkta rol alan hücrelerin hızlı ve etkili biçimde iletişime geçmesi, etkinliklerinin artıtılması ve çeşitli hücrelerin uyarılmasını sağlayan proteinlerdir. İmmün sistem hücreleri arasında iletişimi sağlayan moleküller sitokinlerdir. Gerçekte sitokinler arasında yer alan kemokinler immün hücrelerin bir organdan diğerine, ya da özellikle bir organ içerisinde bir yerden diğerine hareket etmelerini sağlarlar(lökosit migrasyonunu düzenleyen ve uyaran çeşitli hücre tipleri üretir. )
- Slides: 51