DNA ve RNAnn Molekler Yaps ve Analizi Genetik


, hücrelerdeki DNA içeriğinin büyük kısmının nukleusta bulunduğunun sitolojik olarak gösterilmesi.")
. Kalıtsal madde kavramına ilişkin ilk önemli çalışma: Pnömokok tipi bakterilerin (Streptococcus pneumoniae)")


 Avery (1944) ve ark.")
. DNA’nın kalıtsal molekül görevini yaptığını ve virusların kalıtsal molekülünün de")

 tarafından DNA’nın molekül yapısının (çift yapısının) açıklanması. sarmal DNA’nın çift")



 DNA (deoksiriboz), RNA (riboz)")



 nukleotit (şeker+N’lu baz+fosfat grup)")


 • İki zincir birbirine")
")


, Primidin Timin (T) yerine")



 DNA’nın yarı koruyucu tipteki replikasyonunun bakteri hücrelerinde kanıtlanması")
 ilk kez ökaryotik")




. E. coli’ den elde edilen, saflaştırılıp tanımlanan,")





 Proteinler Replikasyon öncesinde, çözülme bölgesinde tek iplikli DNA’yı sabitleştirme, çözülmüş")


 RNaz H aktivitesi yok eder")
. Her biri çoklu")

 replikasyonun Başlatıcının 9 -mer diziye bağlanması ve 13 -mer dizi")







 S evresinde her kromozomun tam")



 Yönetir. Replikatörün seçimi pre-RC’ler’in oluşumuyla yapılır. pre-RC’nin oluşumuna")



 uçların replikasyonu replikasyon çatalında tam olarak yapılamaz.")




 Nukleotid eklemek için bir kalıba gereksinimi olması. (2)")
 Telomerde")








 süreci, aşamaları ve yer alan biyokimyasal moleküller ? ? ?")
 Kalıp bağlama: Bakterilerde RNA polymerazın sigma")

 Konsensus dizileri: , Çeşitli genlerde RNA polimerazın etkin")
’de m. RNA sentezi RNA pol. III kor enzimi faktör sigma Promotor")
 baslangıcı Kalıp zincir Ortak zincir ile m. RNA zinciri aynıdır")
 baslangıcı 3’ 5’ 5’ 3’ 5’ yeni m. RNA zinciri")


 prokaryotlara göre daha karmaşıktır Çekirdekte başlar 3 ayrı RNA")
 • • • Transkr. başlaması için 5’ a bağlı“etki")
 m. RNA’nın işlenmesi gerekir (post-transcriptional ) Pre-m. RNA -------")
 m. RNA’nın işlenmesi gerekir (post-transcriptional ) Pre-m. RNA -------")
 m. RNA’nın işlenmesi gerekir (post-transcriptional ) • Splicing (intron")

 Splicing; Pre-m. RNA olgun m. RNA")

 Organizmalardaki proteinlerin birincil yapılarının (amino asit dizilerinin) DNA molekülündeki genlerin nükleotid")


 3 kodon (UAA, UAG, UGA)")
: aynı anlamı veren birden fazla kodonun bulunması. bir proteinin amino")





- Slides: 113
DNA ve RNA’nın Moleküler Yapısı ve Analizi
Genetik Materyalin özellikleri • • Kendini eşleme Bilgi depolama Bu bilgiyi ifade etme Mutasyon ile çeşitlendirme
Feulgen (1937), hücrelerdeki DNA içeriğinin büyük kısmının nukleusta bulunduğunun sitolojik olarak gösterilmesi.
Griffith (1928). Kalıtsal madde kavramına ilişkin ilk önemli çalışma: Pnömokok tipi bakterilerin (Streptococcus pneumoniae) (hastalık yapma ile ilgili) bazı özelliklerinin bu özellikleri taşımayanlara geçebildiğinin gösterilmesi (transformasyon) Farelerde pnömoniye yol açan ve polisakkarit kapsülle çevrili S ırkı ile hastalık yapmayan ve kapsülsüz R ırkı
canlı R ırkı fare canlı S ırkı fare hastalık ve ölüm yüksek sıcaklık etkisiyle öldürülmüş S ırkı fare canlı ölü S ırkı + canlı R ırkı fare hastalık ve ölüm !!!!
Griffith'in deneyleri kalıtsal maddenin varlığının biyolojik olarak ilk kez gösterilmesi ÖNEMLİ transformasyona neden olan maddenin (kalıtsal molekülün) yapısı ? ? ? proteinler ! ? ! (1930’larda) DNA timonükleik asit (timus nükleik asiti) (sadece hayvanlarda !!!) RNA maya nükleik asidi (sadece bitkilerde !!!)
Kalıtsal Molekül Olarak Nükleik Asitler ve Moleküler Genetiğin Gelişmesi (19441973) Avery (1944) ve ark. Transformasyon deneyinin S tipi pnömokoklara ait saf DNA kullanılarak tekrarlanması Transformasyona neden olan (kalıtsal molekül) madde kesin olarak 1944 yılı moleküler genetiğin başlangıcı D N A
Hershey ve Chase (1952). DNA’nın kalıtsal molekül görevini yaptığını ve virusların kalıtsal molekülünün de nükleik asitler olduğunu gösteren deneysel bir kanıt: Escherichia coli'nin T 2 bakteriyofajıyla bulaştığında bakteri içine giren ve çoğalarak yeni fajları meydana getiren kısmın DNA olduğunun gösterilmesi. DNA’nın kalıtsal molekül olduğunu kesin biçimde kanıtlayan önemli aşama
T 2 fajının morfolojik yapısı Fajın konak bakteri içerisinde çoğalması
Watson ve Crick (1953) tarafından DNA’nın molekül yapısının (çift yapısının) açıklanması. sarmal DNA’nın çift sarmal yapı modeli, molekülün yapısal özelliklerinin ve işlevlerinin açıklanmasında çok önemli bir temel taşı ! ! ! (1) DNA hücrenin fenotipine ait genetik bilgiyi taşır. DNA’daki bilgilerin önce RNA biçiminde kopyası çıkarılır ve bu kopya daha sonra protein biçimine çevrilir. Genetik bilginin akış yönü ana kural (sentral dogma) (2) DNA kendine benzerini yaparak çoğalabilir, hücrenin kalıtsal molekül miktarının iki katına yükselmesinde kalıp görevi yapar.
Watson&Crick, DNA’nın ikili sarmal yapı modeli
Kalıtım Materyali • DNA – Prokoryatlar; bakteriler, arkealar – Okoryatlar; maya, mantar, bitkiler, memeliler • RNA – TVM (tütün mozaik virusu) – Retrovirusler (HIV) • Prion (bakteri ve viruslerde)
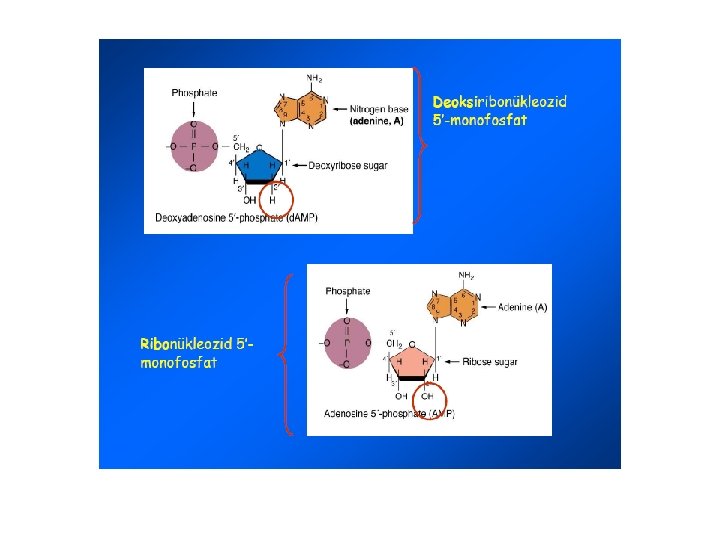
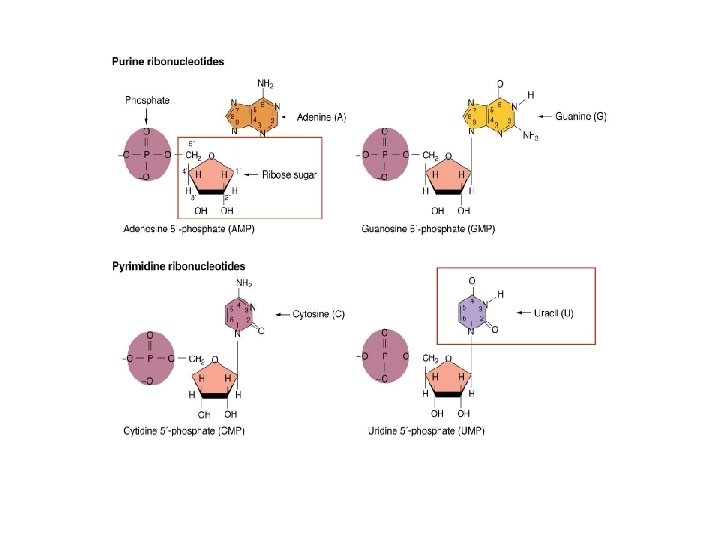
Nukleik asitlerin moleküler yapısı • İşlevi; bütün hucreler için kalitim bilgisi taşırlar • 5 C’lu şeker (Pentoz) – DNA (deoksiriboz), RNA (riboz) • N’lu baz – 9 atomlu-2 halkalı Purinler (A, G) – 6 atomlu-tek halkalı Primidinler (C, T, U) • Fosfat grubu
5 C’lu şeker (Pentoz) DNA (deoksiriboz), RNA (riboz)
N’lu bazlar 9 atomlu-2 halkalı 6 atomlu-tek halkalı
N’lu bazlar • Purinler; A, G – 5 C’lu şekerin C-1 atomuna N-9 atomu ile kovalent bag yaparak baglanirlar • Primidinler; T, C, U – 5 C’lu şekerin C-1 atomuna N-6 atomu ile kovalent bag yaparak baglanirlar
Fosfat grubu • 5 C’lu sekerin C-3 ve C-5 atomu ile baglanarak, 3’-5’ fosfodiester baglari olusturular
nukleozit (şeker+N’lu baz) nukleotit (şeker+N’lu baz+fosfat grup)
nukleotitler • 3’-5’ fosfodiester baglari ile nukleik asitlerin temel yapisini olusturular – dinukleotit; iki mononukleotit 3’-5’fosfodiester baglari ile baglanirlar – oligonukleotit; 20 yada daha az sayida nukleotit iceren zincir – polinukleotit; uzun zincirler
Watson-Crick DNA Modeli
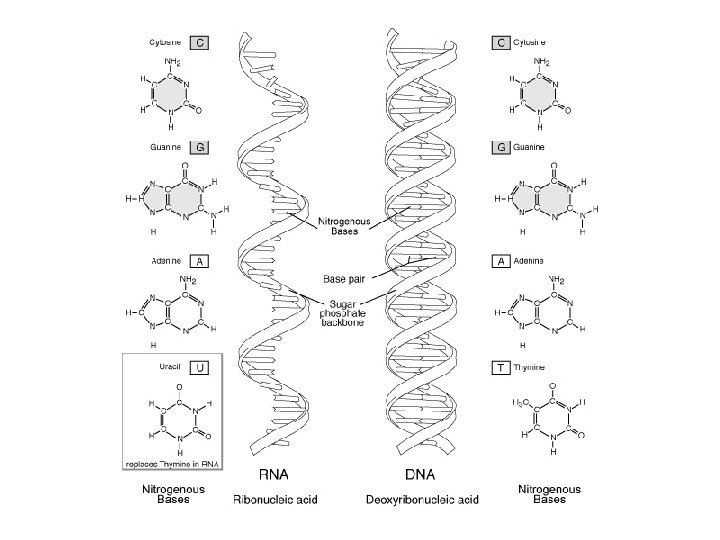
Watson-Crick DNA Modeli • Sağ el yönlü-çift sarmaldır (double helix) • İki zincir birbirine antiparaleldir – (5’-3’ ve 3’-5’ yönü) • N’lu bazlar eşlenik (complementary) olarak, zayıf H bağları ile bağlıdır A ile T (2 H bağı) G ile C (3 H bağı)
Watson-Crick DNA Modeli • N’lu bazlar arası mesafe 3. 4 A° (0. 34 nm) dir • Sarmalın tam bir dönüşü 34 A ° (3. 4 nm) 10 baz yer alır • Sarmal ekseni üzerinde büyük ve küçük oluklar yer alır • Sarmalın çapı 20 A ° (2 nm) • B formundadır
X-ışını kırımı’na göre DNA Formları • A-DNA (sağ-el sarmal, çapı: 23 A°, tam dönüş: 11 baz, BDNA’ya göre daha sıkı yapıda) • B-DNA (sağ-el sarmal, çapı: 20 A°, tam dönüş: 10 baz) • Z-DNA (sol-el sarmal, çapı: 18 A°, tam dönüş: 12 baz, sıkı ve zig-zag yapıda) • C-DNA (sağ-el sarmal, çapı: 19 A°, tam dönüş: 9. 3 baz, BDNA’ya göre daha az sıkı yapıda) • D, E-DNA (sağ-el sarmal, G yoktur, tam dönüş: 7. 5 -8 baz, BDNA’ya göre daha sıkı yapıda) • P-DNA (sağ-el sarmal, B-DNA’ya göre ince, uzun yapıda, tam dönüş: 2. 62 baz)
A B Z -DNA
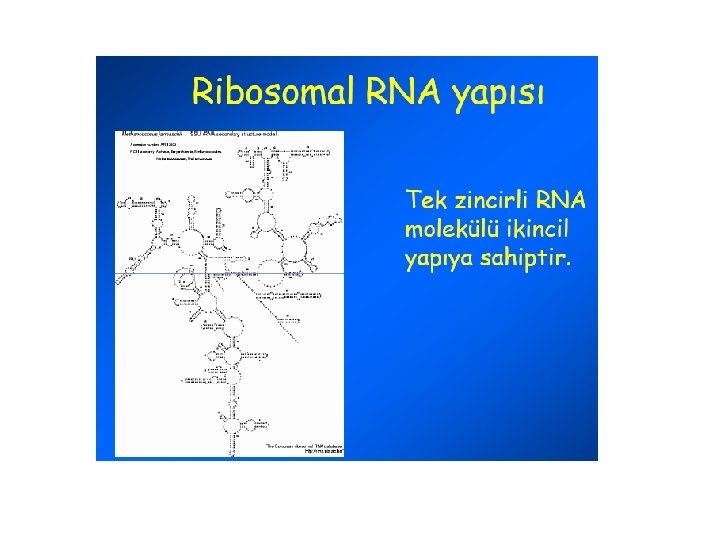
RNA • Tek zincirli polinukleotitdir • Pentoz; (5 C’lu Riboz), Primidin Timin (T) yerine Urasil; (U) bulunur • Formları; m. RNA, t. RNA, r. RNA, sn. RNA • Çekirdekçik, stoplazma ve ribozomlarda bulunur
Nukleik asitlerin Denaturasyonu ve Renaturasyonu • Deneturasyon; H bağları kopar, çift yapı çözünür, zincirler birbirinden ayrılır – Isı ve kimyasal yolla olur
REPLİKASYON
Kalıtsal molekül olan nükleik asit moleküllerinin kendine benzer moleküller meydana getirerek çoğalmaları. Böylece hücredeki kalıtsal molekül miktarı iki katına yükselir. Amacı? ? • genetik bilginin dölden döle devamlılığı Nasıl? ? ? • Hücre bölünmesi • kalıtsal molekülün eşit biçimde paylaşımı • yavru hücrelerin birbirleriyle ve ana hücreyle aynı miktarda kalıtsal moleküle sahip olmaları
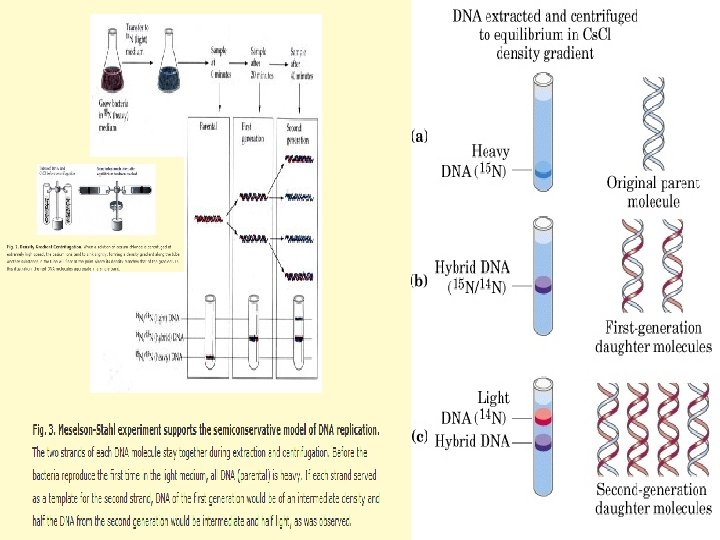
Replikasyon modelleri Meselson ve Stahl (1958) DNA’nın yarı koruyucu tipteki replikasyonunun bakteri hücrelerinde kanıtlanması
• J. H. Taylor, P. Woods, and W. Hughes (1957) ilk kez ökaryotik DNA sının replikasyonunun yarı korunumlu olduğunu otoradyografik yöntemle vicia faba ‘da gösterdiler. • (3 H- timidinli )
Replikasyon orijini; eşlemenin başladığı yerdir Replikasyon çatalı: Kromozom üzerinde replikasyon olduğu noktada sarmala ait zincirlerin açılması ile ortaya çıkan çatala rep. çatalı denir. Replikon: bir orijinden replikasyon başladıktan sonra replike olan DNA’nın uzunluğunun birim olduğunu belirtir. • Tek yönlü, çift yönlü • çift yönlü eşlemede; orijinden itibaren zıt yöne doğru ilerleyen iki replikasyon çatalı oluşacaktır
Cairns, • E. Coli de radyoizotop kullanarak otoradyografi yöntemi ile DNA replikasyonunu izlemiş ve replikasyonun bir noktadan başladığını gözlemlemiştir. Ori. C olarak adlandırılan bu özgül bölgenin konumu E. coli kromozomu üzerinde haritalanmıştır. 254 bç’lik bir bölgedir (9 mer ve 13 mer tekrar dizileri). Bu bölgenin çok az kısmı DNA sentezinin başlaması için gereklidir • • Bakteriyofaj ve bakterilerde DNA sentezi bir noktadan başladığı için, kromozomun tümü bir replikondur. Daha sonraki araştırmalarda replikasyon iki yönlüdür. Başlangıç ori. C her iki yönünde hareket eder (5’-3’ çift yönlü- yarı-saklı DNA rep. ) • Dolayısıyla iki replikasyon çatalı oluşurken ter olarak adlandırılan sonlanma bölgesinde birbirleriyle birleşir.
REPLİKASYONDAKİ ENZİM SİSTEMİ DNA polimerazların kataliz aktiviteleri 5' 3' polimeraz aktivitesi : replikasyondaki her reaksiyon aşamasında bir nukleotidin 3’OH grubu ile eklendiği ipliğin ucundaki (son) nukleotidin 5’PO 4 grubu arasında fosfodiester bağı oluşturulması sentezlenen DNA ipliğinin 5' 3' yönünde uzaması. 3' 5' eksonukleaz aktivitesi : replikasyon sırasında sentez edilen zincirin yapısına yanlış giren nükleotidin yok edilmesi. 5' 3' eksonukleaz aktivitesi: replikasyonda görevi tamamlanmış olan primer zincirin yok edilmesi; ayrıca DNA’da kusurlu bölgelerin kesilmesi
DNA polimerazın hata giderici 3’ 5’ eksonukleaz aktivitesi uzamakta olan DNA’nın 3’OH ucunda etkili
Bakterilerdeki DNA Polimerazları Kornberg ve arkadaşları (1957). E. coli’ den elde edilen, saflaştırılıp tanımlanan, replikasyonla ilgili enzim DNA polimeraz I (Kornberg enzimi) Bakterilerde (E. coli’de) en az 5 farklı DNA polimeraz (DNA polimeraz I-V).
Özet olarak Polimeraz I-II-III in vivo rollerine bakılırsa; Polimeraz I: Primeri uzaklaştırır ve primerler uzaklaştıkça doğal olarak oluşan boşluklarda DNA sentezleyerek bu boşlukları doldurur. Ekzonükleaz aktivitesi ile bu işlem esnasında oluşabilecek hatalarıda onarır. Kısaca 5' - >3' DNA polymerase aktivitesi 3' ->5' ekzonukleaz aktivitesi (hata okuma mekanızması= proofreading) 5' - >3' ekzonukleaz aktivitesi (DNA tamiri sırasında oluşan boşlukların doldurulması = nick translation) Polimeraz II: Ultraviyole gibi dış etmenler sonucu hasar gören DNA ‘nın bozulduğunda onarımında rol alır. Enzim replikasyon çatalındaki DNA sentezi bozulduğunda aktive olan bir gen tarafından şifrelenmektedir. , 90 k. D mol. ağ. 5' - >3' DNA polymerase aktivitesi 3' ->5' ekzonukleaz aktivitesi (hata okuma mekanızması= proofreading) Polimeraz III: Replikasyondan asıl sorumlu olan enzimdir. Enzimin 3’-5’ ekzonükleaz aktivitesi ile sentez sırasında hata onarım işlevini görmesini sağlamaktadır.
Polimerazlar I II III Zincir sentezinin başlatılması - - - 5’-3’ yönlü polimerizasyon + + + 3’-5’ yönlü ekzonukleaz aktivitesi + (hata okuma mekanızması= proofreading) + + 5’-3’ yönlü ekzonukleaz aktivitesi + (DNA tamiri sırasında oluşan boşlukların doldurulması = nick translation)
Replikasyon sırasındaki karmaşık olayların çözülmesi gerekmektedir. Sentezin 5’-3’ yönünde replikasyon çatalı oluşturarak gerçekleştiği bilinmektedir. Bu replikasyon çatalının 2 zıt yöne doğru hareket etmektedir. Bu noktada bazı noktalar aydınlatılmalıdır. a) b) c) d) e) f) g) Her iki zincirde sentezin devamı için ‘açık’ konfigürasyonun dayanıklı olmasını sağlayan bir mekanizma gerekir Sarmalin açılması ve zincirin daha aşağı kısımlarda tekrar sarılması sonucu ortaya çıkan gerilimi azaltmak için bir mekanizma bulunmalıdır Pol III’ün 5’-3’ yönlü polmerizasyonu için primerlere (RNA primerler) gereksinim vardır. 5’-3’ ucu için sorun olamasa bile 3’-5’ ucu pol. III için sorun oluşturur ve bu zincirdeki sentez zıt yönde ve kesintilidir Rep. tamamlanmadan önce RNA primerler uzaklaştırılmalıdır. Boşlukları doldurmak için yeni sentezlenen DNA bitişiğindeki DNA zinciri birleştirilmelidir Kopyalama sırasında DNA polimerazlar eşlenik bazları doğru takmaktadırlar. Hatalı takılan bazlar bir hata okuma mekanizması (proofreading) tarafından düzetilmelidir
REPLİKASYONDA GÖREV ALAN DİĞER ENZİMLER VE YAPILAR
HELİKAZ DNA helikazlar, replikasyon çatalının ilerlemesi için çift sarmalı çözen enzimler; halka biçiminde heksamerik proteinlerdir. Tek iplikli DNA’ya bağlanıp, ATP hidrolizi enerjisini kullanarak, DNA zinciri boyunca hareket edip diğer DNA zinciriyle bağlantısını yok ederler.
Tek Zincire Bağlanan (SSB) Proteinler Replikasyon öncesinde, çözülme bölgesinde tek iplikli DNA’yı sabitleştirme, çözülmüş bölgenin kalıp olarak iş görebilmesinde görev alırlar. DNA helikaz çözülmeyi sağladıktan sonra, SSB proteinler tek ipliklere hızla bağlanırlar. Bir SSB proteinin zincire bağlanması başka bir SSB’nin hemen yanına bağlanmasını teşvik eder (işbirlikçi bağlanma) SSB proteinlerle kuşatılan DNA kalıp olarak kullanılabilir. SSB’lerin bağlanmalarında dizi özgüllüğü yoktur; DNA molekülünün şeker-fosfat omurgası ve bazlarıyla elektrostatik etkileşimle ilişki kurarlar.
DNA topoizomerazlar içinde yer alan DNA giraz enzimi Çözülme sırasında kapalı molekülde negatif dönümlerin azalmasıyla oluşan (ve çatalın ilerlemesinde engelleyici etki yapan) pozitif süper dönümlerin yok edilmesi. Halka biçimindeki DNA’da negatif süper dönümler meydana getirerek replikasyon çatalındaki çözülmenin teşvik edilmesi. Oluşan kırıklar sonra tekrar birleştirilir. Bu çeşit reaksiyonlarda ATP hidrolizinden açığa çıkan enerji kullanılır.
DNA sentezinin başlaması için RNA primerlerine gereksinim vardır Sarmal açıldıktan sonra sentez başlayabilir fakat DNA polimeraz III’ün polinükleotit zincirini uzatması için serbest 3’-OH grubu olan bir primer gereklidir. A) Kalıp DNA üzerinden, DNA’ya eşlenik olan kısa RNA parçası sentezlenir (5 -15 nukleotid uzunluğunda) B) RNA sentezi PRİMAZ adı verilen RNA polimerazın bir çeşiti tarafından katalizlenir. C) Primazın sentezi başlatması için serbest 3’ ucu gerekmez ve DNA poly. III bu RNA parçasını 5’ – deoksiribonükleotitleri takmaya başlayarak Dna sentezine başlatır. D) Daha sonra RNA primeri uzaklaştırılarak DNA kalır. Bu reaksiyon DNA poly 1 ile olur. Bu işlem bakteri, virüs ve çeşitli ökaryatik organizmalarda evrenseldir. Primozom, primer sentezi için gerekli reaksiyonlara katılan protein kompleksi.
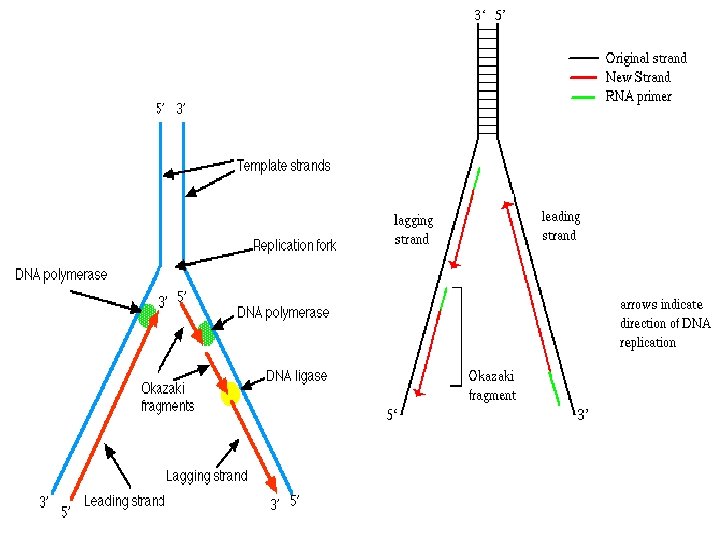
RNA primerlerin çoğunu (DNA ile baz eşleşmesi yapmış RNA’yı) RNaz H aktivitesi yok eder (“H” : RNA: DNA hibridi). DNA’nın ucundaki son ribonukleotid 5’ 3’ eksonukleaz aktivitesiyle ortadan kaldırılır. RNA primerin yok edilmesi, DNA polimeraz için uygun primer: kalıp bağlantı yeri yaratır. DNA polimerazın 3’ ucu kullanılıp boşluk kapatmasından sonra ipliklerin 3’OH ve 5’PO 4 uçları arasında kalan çentik DNA ligaz tarafından kapatılır.
Ökaryotlardaki DNA Polimerazlar. Ökaryotlarda DNA polimeraz çeşidi daha fazla ( 15). Her biri çoklu altbirimlerden oluşur. DNA polimeraz : nukleustaki DNA replikasyonunun başlamasında esas enzim (replikaz). DNA polimeraz /primaz yeni DNA zincirlerinin sentezinin başlatılmasında rol oynayan 4 alt birimli bir protein kompleksi (2 DNA polimeraz : 2 primaz). DNA polimeraz /primaz yavaş ilerleyici tipte; başlatıcı görevini tamamladıktan sonra yerini DNA polimeraz veya DNA polimeraz alır (polimeraz değişimi ). DNA polimeraz : mitokondrilerde (ve kloroplastlarda ? ) DNA replikasyonu. Diğer polimeraz tipleri (özellikle, DNA polimeraz ) onarımda iş görürler.
Bakterilerde replikasyon
Bakterilerde mekanizması: (E. coli’de) replikasyonun Başlatıcının 9 -mer diziye bağlanması ve 13 -mer dizi ile etkileşime girip 20 bç’lik çözülmeye yol açması. DNA helikaz ve helikaz yükleyicinin açığa çıkan tek iplikli DNA kalıbına bağlanması. Helikaz moleküllerinin 5’ 3’ yönünde hareket etmeleri. Helikaz aracılığıyla başlangıç yerine bağlanan DNA primazın her iplikte bir RNA primerini sentezlemesi.
DNA Pol III holoenziminin, primer: kalıp ve helikaz etkileşimleriyle başlangıç yerlerine getirilmesi. RNA primerlerine gelen kayan kelepçelerin önden giden zincirin sentezini yapan polimerazlara yol göstermeleri. Primazın arkadan gelen ipliğe ait ilk primerleri sentezlemesi. Yeni primer: kalıp bağlantılarına, önden giden zincirlerdeki kayan kelepçelerin bağlanması. DNA Pol III tarafından arkadan gelen DNA iplik sentezinin başlatılması.
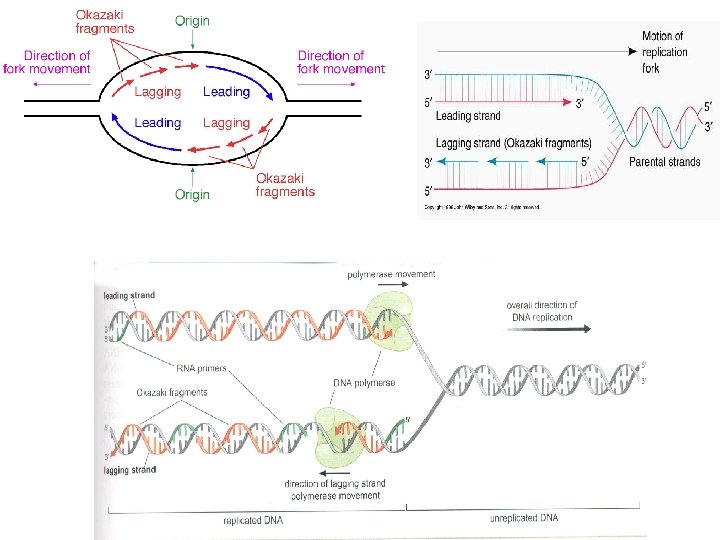
DNA Replikasyonunun Kesintili ve kesintisiz olarak gerçekleşir. DNA sarmalında zincirlerden birisi 5’-3’ iken diğer, 3’-5’ yönünde olduğu durumundadır. Bu durumda DNA Poly. III, DNA sentezini sadece 5’-3’ gerçekleştirebilir. Bu durumda replikasyon çatalı açıldıkça ve aşağı doğru hreket ettikçe sadece bir kol sürekli DNA sentezi yamaktadır. Bu zincire kesintisiz DNA zinciri (leading DNA strand denilmektedir. Diğer DNA zincirinde sentez için pekçok başlangıç noktası gereklidir. Dolayısıyla buda kesintili DNA sentezi yapılır. Okazaki ve ark. (1968) E. Coli de bakteriyofaj DNA sının replikasyonu sırasında yeni sentezlenen DNA’nın kalıp zincire H bağlarıyla tutunan 1. 000 -2. 000 nükleotidlik küçük parçalar halinde bulunduğunu göstermiştir. RNA primeri bu şekildeki herbir parçanın bir kısmını oluşturmaktadır. Okazaki parçaları denilen bu parçacıklar sentez devam ettikçe molekül ağırlığı artacaktır. (ökaryotlarda 100 -400 nukleotid) Kesintili DNA sentezinde, RNA primerlerini uzaklaştıracak ve okazaki fragmanlarını birleştirecek enzimlere gereksinim vardır. Poly I bu işten sorumludur. Bu parçalarda DNA ligaz ile birleştirilir.
Replikasyon çatalında işlev gören tüm proteinlerin oluşturduğu birlik REPLİZOM Replizomdaki proteinlerin eşgüdümü aralarındaki etkileşimlerle sağlanır. Çok sayıda protein-protein etkileşimleri replikasyon çatalının hızlı şekilde ilerlemesi.
Ökaryotlarda DNA Replikasyonu Ökaryotik hücrelerde DNA replikasyonu, prokaryotlardan farklı olarak, hücre döngüsünde sadece belli bir kısmında yapılır.
Hücre döngüsü G 1 replikasyonun yapılmasına hazırlık oluşturan metabolik olaylar. S DNA replikasyonu G 2 mitoz bölünmeye hazırlık oluşturan metabolik olaylar M mitoz bölünme
Ökaryotik hücreler çok fazla miktarda DNA içerirler (örneğin, baklanın kromozomlarında ~800 cm, insandaki büyük kromozomlarda ~7 cm’lik DNA). Her bir kromozomdaki DNA’da çok sayıda replikasyon başlangıç noktası (çok sayıda replikon) bulunur. Replikonlar birbirinden bağımsızdır, replikasyonun başlangıç ve bitimi hepsinde aynı zamanda olmayabilir.
Ökaryotik Kromozomlar Her Hücre döngüsünde Bir Kez Replikasyon Geçirirler Hücre çevriminin S evresinde tüm DNA’nın tam olarak iki katına yükselmesi gerekir. ¨Kromozomun bir kısmının replikasyonunun tamamlanmaması ¨ yavru kromozomlar arasında uygun olmayan bağlantıların meydana gelmesi ¨ kromozom kırıkları veya kayıpları. ¨DNA replikasyonunun tekrarlanması ¨ genomun bazı bölgelerinde kopya sayısının artması ¨ önemli düzenleyici gen kopyalarının artışıyla gen anlatımında, bölünmesinde ya da sinyallere yanıtta bozulmalar. hücre
Çok sayıda replikasyon başlangıç yeri taşıyan ökaryotik kromozomlarda: a) S evresinde her kromozomun tam olarak replikasyon geçirmesi için yeterli sayıda başlangıç yerinin aktif duruma getirilmesi gerekir. b) Bir başlangıç yerinin hücre bölünmesinin bir sonraki çevirimine kadar aktif olmayan durumda kalması gerekir.
• Ökaryotik replikonlarda, prokaryotlardaki gibi, zıt yönlü iki replikasyon çatalı oluşur. • Replikasyon, iki iplikte birden aynı anda ve iki yönlü (yarı-kesintili biçimde) yapılır. • Ökaryotik replikonların boyları prokaryotlardakinden çok kısa ve replikasyon hızları daha azdır
Ökaryotlarda DNA’nın kromatin yapısında oluşu, nedeniyle replikasyonun başlaması için DNA-histon kompleksinin yok olması, replikasyonla oluşan yeni DNA moleküllerinin de kromatin yapısını kazanmaları gerekir (olayı karmaşıklaştırıcı etkiler !!!)
Ökaryotlarda Replikasyonun Başlaması Replikasyonun başlaması hücre çevriminde farklı zamanlarda meydana gelen iki aşamayı gerektirir: Replikatörün seçimi, G 1 evresinde replikasyonun başlamasını yöneten dizilerinin belirlenmesidir. Genomdaki her replikatörde çoklu protein kompleksinin bir araya gelmesine neden olur. Başlangıç yerinin aktifleştirilmesi, S evresinde DNA’nın çözülmesinin ve DNA polimerazın bağlanmasını başlatmak üzere replikatörle ilişkili protein kompleksinin tetiklenmesidir. Bu süreçlerin zamansal farklılığı her hücre döngüsünde her kromozomun ancak bir kez replikasyon geçirmesini sağlar.
Replikasyonun başlamasını “Replikasyon Öncesi Kompleks” (pre-RC) Yönetir. Replikatörün seçimi pre-RC’ler’in oluşumuyla yapılır. pre-RC’nin oluşumuna katılan proteinler: “Başlangıç Tanıma Kompleksi” (“origin recognition complex”, ORC) altı proteinlik bir kompleks, replikatörü tanıyıp bağlanır. Helikaz yükleyiciler (Cdc 6 ve Cdt 1), ORC’nin bağlanmasıyla, o bölgede toplanır. Helikaz (Mcm 2 -7 kompleksi) replikasyon çatalına bağlanır.
Başlangıç yerinin aktifleştirilmesi G 1’de oluşan pre-RC’ler S evresinde replikasyonu başlatmak üzere aktifleşirler: pre-RC’ler iki protein kinaz (Cdk ve Ddk) tarafından aktif duruma getirilir. Kinazlar S evresine girildiğinde pre-RC ile diğer replikasyon proteinlerini fosforiller. Diğer replikasyon proteinleri (üç DNA polimeraz, bunları bir araya getiren Mcm kompleksi, Cdc 6 ve Cdt 1 vb) başlangıç yerinde bir araya gelirler ve replikasyon başlar. DNA polimerazların bağlanması için gerekli faktörler görevleri tamamlandıktan sonra serbest kalır ya da bozulurlar.
Ökaryotik hücrelerde çok sayıda replikasyon başlangıcının her hücre çevriminde bir kez aktif duruma geçmeleri, pre-RC’lerin oluşumunun ve aktifleşmesinin sikline bağımlı kinazlar (Cdk’lar) tarafından sıkı şekilde kontroluyla sağlanır. Cdk’lar pre-RC’lerin işlevinin düzenlenmesinde iki zıt rol oynar: (1) DNA replikasyonunu başlatmak için pre-RC’leri aktifleştirirler. (2) Yeni pre-RC’lerin oluşumunu engellerler.
Ökaryotlarda replikasyonun moleküler mekanizması
Replikasyonun Tamamlanması Ökaryotlardaki doğrusal biçimli kromozomlarda (DNA’larda) uçların replikasyonu replikasyon çatalında tam olarak yapılamaz. Arkadan gelen ipliklerde, Okazaki parçalarının sentezi için gerekli son RNA primer ortadan kaldırıldığında kromozomun ucunda replikasyon geçirmemiş kısa tek iplikli bir DNA bölgesi kalır uç replikasyonu sorunu.
Her replikasyon çevriminde iki yavru kromozomdan birinin boyunda kısalma olması ve dölden döle geçerken kromozomların uçlarındaki genler kaybolması sorunu !!!
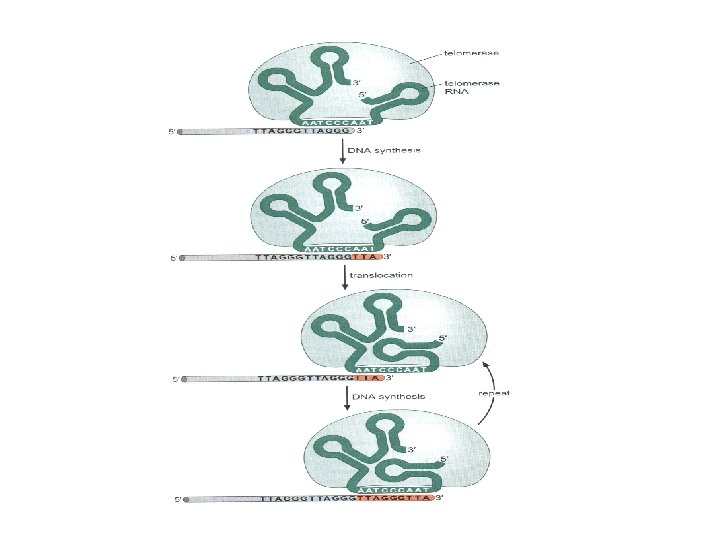
Ökaryotlarda uç replikasyonu sorununu ortadan kaldırmak için geliştirilmiş özel mekanizma: Kromozomun 3’ ucunun telomeraz enzimi tarafından uzatılması • Ökaryotik kromozomların uçlarında (telomer) çok sayıda (30 -70 kez) tekrarlanan TG’ce zengin DNA dizileri bulunur (örneğin, insanlarda 5’-TTAGGG-3’). • Tekrarların bir kısmı, kromozomun ucundan dışarı doğru tek zincirli DNA şeklinde 3’ yönünde uzamış durumda. • Uzamış uç yeni bir replikasyon başlangıç yeri olarak iş görür ve özel bir DNA polimerazın (telomeraz) o bölgede toplanmasını sağlar.
Telomeraz = protein + RNA d. NTP’lerin eklenmesi için kendi RNA’sını kalıp olarak kullanarak özel tek zincirli DNA dizilerinin 3’ ucunu uzatır. RNA’da telomer dizinin tamamlayıcısı olan nukleotid dizisi bulunur. Eşleşme, RNA kalıbının bir kısmını tek zincirli durumda tutacak (bir primer: kalıp bağlantı yeri yaratacak) şekilde meydana gelir. Protein kısmı ters transkriptaz enzimlerine benzer. RNA kalıbının ucuna DNA’yı ekler fakat RNA’yı kopyalamayı fazla devam ettirmez. Kalıp RNA DNA ürününden çözünür, telomerin son üç nukleotidiyle yeniden eşleşir ve bu süreç tekrarlanır.
Telomeraz telomerin 3’ ucunu kullanıp TG’ce zengin dizileri ekleyip ipliği uzatır. Uzatılmış 3’ uç, arkadan gelen zincirdeki replikasyon aygıtının DNA’nın 5’ ucunu uzatabilmesi için ek kalıp sağlar. Primaz ve DNA polimeraz bu kalıptan yararlanarak tamamlayıcı dizileri sentezlerler. Kromozomun ucunda tek zincirli bir DNA bölgesi kalmakla birlikte, telomerazın etkisi ve arkadan gelen zincirdeki replikasyon aygıtı telomeri yeterli uzunlukta tutar. kromozomun uç kısmı kısalmaktan korunur.
Telomerazın diğer DNA polimerazlara benzerlikleri: (1) Nukleotid eklemek için bir kalıba gereksinimi olması. (2) Sadece DNA’nın 3’ ucundan uzatma yapabilmesi. (3) Aynı nukleotid öncüllerini kullanması. (4) İlerleyici tipte olması. Telomerazın diğer DNA polimerazlardan farkları: (1) (2) (3) (4) RNA bileşeni taşıması. Dışarıdan bir kalıp gereksinimi olmaması. Tek zincirli DNA kalıbı kullanma yeteneği. RNA-DNA helikaz aktivitesi içermesi.
Telomerin boyunun düzenlenmesi: Telomerin çift zincirli bölgesine bağlanan proteinler (telomeraz aktivitesinin zayıf engelleyicileri) Telomerde tekrarlanan dizilerin kopya sayıları az olduğunda az sayıda protein telomere bağlanır telomerazın aktivitesi artırılır. Telomerler fazla uzun olduğunda proteinler birikir telomerazın aktivitesi engellenir.
Mitokondri DNA’sının Replikasyonu Yer değiştirme tipindeki replikasyon mekanizması: Başlangıçta sadece bir ipliğin (memelilerde hafif iplik) tamamlayıcısının sentezi yapılmaya başlar; replikasyon çatalı halkasal DNA’nın yaklaşık yarısına geldiğinde ise senteze diğer iplikte (memelilerde ağır iplik) devam edilir. iki iplikteki sentez farklı zamanda tamamlanır.
TRANSKRİPSİYON
Transkripsiyon: DNA’daki bilginin RNA’ya aktarılması • RNA – Tek zincirli bir polinukleotid – Riboz seker icerir – DNA’daki timin yerine urasil icerir – Olgunlaşmış m. RNA molekullerinin 5’ ve 3’ uçları işlevsel önem taşır.
• RNA esleniklik kuralına uygun olarak kalıp DNA zinciri uzerinden transkribe olur. • Genlerin buyuk kısmı proteinleri kodlarken az bir kısım gen ise ifadesi olmayan islevsel RNA’ları kodlar. • Genin nukleotid dizilimi bir proteindeki aminoasitlerin sırasını belirler. Bu dizilim proteinin seklini, boyutunu ve islevini belirler. • m. RNA uclu nukleotid grupları (kodon) halinde t. RNA antikodonunun m. RNA kodonu ile eslesmesi aracılığı ile ribozomlarda tercüme (ifade) edilir.
RNA cesitleri m. RNA: Protein kodlayan – Baslıca transkriptlerdir – Okaryotlarda transkripsiyon sonrası islemden gecirilir • 5’ ve 3’ uc modifikasyonu • Intronların uzaklastırılması • Islevsel/Yapısal RNA t. RNA: amino asitleri ribozoma tasır. r. RNA: ribozomların yapısal ve katalitik bilesenleridir. sn. RNA: spliceosome’un yapısal ve katalitik bilesenleridir. sno. RNA: kucuk nukleolar RNA r. RNA olgunlasmasına katılır. sc. RNA: sitoplazmadaki protein trafiğini yonlendirir. si. RNA: small interfering RNA gen ifadesinin kontrolunde rol alır.
Transkripsiyon • RNA polimeraz RNA sentezini katalizler Transkripsiyon asimetriktir – DNA zincirlerinden birini kalıp olarak kullanır. RNA transkripti kalıp olmayan zincirle aynı dizilime sahiptir. • Belirli bir gen icin hep aynı zincir uzerinden gerceklesir. – DNA ifade edilecek kısmı ile acılır – Uzayan RNA zincirinin 3’ ucuna serbest nukleotidleri takar. • 5’ den 3’ yonune RNA sentezi. (3’ ve 5’ yönünde okunur)
RNA polimeraz • Prokaryotlar: tek bir RNA polimeraz – m. RNA, r. RNA and t. RNA – Trankripsiyon ve translasyon birbiri ile esli gider • Okaryotlar: uc RNA polimeraz – RNA polimeraz I: r. RNA genleri – RNA polimeraz II: m. RNA Transkriptler islenir – RNA polimeraz III: t. RNA, 5 S r. RNA • Okaryotlarda transkripsiyon cekirdekte translasyon sitoplazmada ribozomlarda gerceklesir.
TRANSKRİPSİYON BASAMAKLARI • Baslangıc – Genin 5’ ucundan – RNA polimeraz promotora bağlanır. – DNA cift zinciri ayrılır • Uzama – Nukleotidler 3’ uctan ilave olur. – GTP’den enerjisi sağlar • Sonlanma – Genin 3’ ucunda gerceklesir. Kodlayıcı bölge 5’UTR -----------3’UTR
m. RNA sentezi (transkripsiyon) süreci, aşamaları ve yer alan biyokimyasal moleküller ? ? ? • Transkripsiyon için gerekli bileşenler – Eşlenik DNA, transkripsiyon faktörleri a)prokartotlarda; factor sigma, b)okaryotlarda TFIIA, TFIIB) - RNA polimerazlar - ribonukleotitler
Kalıba bağlanma, promotorlar ve sigma alt faktörü 1) Kalıp bağlama: Bakterilerde RNA polymerazın sigma alt biriminin promotor denilen özgül DNA dizilerini tanımasıyla gerçekleşir. Bu bölgeler genin transkripsiyonunun başlangıç noktasına göre daha yukarıda 5’ bölgede yer almaktadır. Enzimin, promotor bölgeyi tanıyana kadar DNA üzerinde belli bir uzunluk ölçüsünde keşif yaptığı ve sarmal 40 nük. transkripsiyonun başlangıç noktasından yukarıda yer alan 60 nük. bölgeye bağlandığı düşülmektedir.
• Enzim bu noktada bağlandıktan sonra, sarmal bu bölgede denatüre olur yani açılır ve DNA kalıbı enzimin çalışması için açık duruma gelir. • Promotorların önemi: Transkripsiyonun başlama etkinliğini idare ederler. Bakterilerde transkripsiyonun her 1 -2 saniyede birden her 10 -20 dk. Bire kadar değişen oranlarda başlamasına neden olan hem güçlü hemde zayıf promotorlar vardır. Mutasyon etkinliği düşürür.
• Promotor-enzim etkileşimi ; 1) Konsensus dizileri: , Çeşitli genlerde RNA polimerazın etkin bağlanmasını sağlayan benzer diziler. Bunlar evrim sürecinde korunmuş olması bunların rollerinin nekadar önemli olduğunu göstermektedir.
Prokaryotlar (E. coli)’de m. RNA sentezi RNA pol. III kor enzimi faktör sigma Promotor bölge: Prokaryotda: 10 bç TATAAT (pribnov kutusu), -35 bç TTGACA (- 35 bölgesi) Okaryorlarda: CAAT (-80) ve TATA (-30) kutu bölgeleri faktör sigma: RNA pol. III’ün DNA promotor bölgesine bağlanarak polimerizasyonun daha kararlı yapıda tutulmasını sağlar
m. RNA sentezi (transkripsiyon) baslangıcı Kalıp zincir Ortak zincir ile m. RNA zinciri aynıdır
m. RNA sentezi (transkripsiyon) baslangıcı 3’ 5’ 5’ 3’ 5’ yeni m. RNA zinciri (5’-3’ yönünde sentezlenir)
m. RNA zincir uzanımı ve bitimi Uzanım: RNA Polimeraz ribonukleotidleri 3’-5’ yönündeki Polimeraz aktivitesi ile 5’->3’ yönünde sentezler Bitim: “dur” (stop) sinyalleri alana kadar olur. ; (durdurma dizisi yaklaşık 40 bç uzunluğundadır) !!!! Prokaryotlarda bu dizi sonderece önemlidir, çünkü bu organzimalarda, bir genin sonu ile başlama noktaları birbirine çok yakındır. Sentez sonucu (öncül) pre-m. RNA’lar oluşur ve olgun (mature) m. RNA haline donusmesi gerekir Prokaryotlarda; sentez sonrası pre-m. RNA= olgun m. RNA Okaryotlarda ise pre-m. RNA, olgun m. RNA değildir pre-m. RNA’nın olgun m. RNA’ya çevrilmesi gerekir
Bazı durumlarda sentezin sonlanması sonlanma faktörü adı verilen rho bağlıdır. Rho uzayan RNA kopyası ile fiziksel ilişki kuran büyük bir hekzomerik proteindir. Sonlanma noktasında, RNA kopyası DNA kalıbını terkeder ve kor polimeraz enzimi de kalıptan ayrılır. Sentezlenen RNA molekülü, genin kalıp zincirini temsil eden DNA dizisine tamamen eşleniktir. Kalıp zincirde, nerede A, T, C yada G varsa, RNA molekülüne sırası ile, U, A, G, C nükleotidleri girmiştir.
ökaryotlarda m. RNA sentezi (transkripsiyon) prokaryotlara göre daha karmaşıktır Çekirdekte başlar 3 ayrı RNA polimeraz tarafından yönlendirilir Tip Ürün Yerleşimi I r. RNA Nukleolus II m. RNA, sn. RNA Nukleoplasma III 5 S r. RNA, t. RNA Nukleoplasma
ökaryotlarda m. RNA sentezi (transkripsiyon) • • • Transkr. başlaması için 5’ a bağlı“etki artırıcılar (enhancers)” gerekir Okaryotik m. RNA islenmesi • 5’ uc: sapka/capping – 5’ uca 7 -methylguanosine ilavesi • 3’ uc: poly(A) kuyruğu – 3’uca 150 -200 adenin nukleotidi eklenir – cis-acting elements (kor promoter) • CAAT (-100) ve TATA (-30) kutusu -trans-acting elements (Polimeaz II bağlanma ) – Transkripsiyon faktörleri: TATA bölgesine bağlanırlar – TBP: (TFIIA), TFIIB)
ökaryotlarda m. RNA sentezi (transkripsiyon) m. RNA’nın işlenmesi gerekir (post-transcriptional ) Pre-m. RNA ------- m. RNA • 5’capping ve 3’ tailing • Splicing (intron bölgeleri atılır, ekzon bölgeler kalır)
ökaryotlarda m. RNA sentezi (transkripsiyon) m. RNA’nın işlenmesi gerekir (post-transcriptional ) Pre-m. RNA ------- m. RNA • 5’capping: 5’ ucuna 7 -metilguanozin eklenir; methilizasyon • 3’ tailing; 3’ ucuna poliadenozinalar tarafından A’lar eklenir. AAAAAA
ökaryotlarda m. RNA sentezi (transkripsiyon) m. RNA’nın işlenmesi gerekir (post-transcriptional ) • Splicing (intron bölgeleri atılır, ekzon bölgeler kalır) – sn. RNAs ve splicing enzimleri – Spliceosome= sn. RNAs + pre-m. RNA • Intron: İşlevsel olmayan (a. a kodlamayan) bölgeler • Ekzon: İşlevsel olan (a. a kodlayan) bölgeler
splicing
ökaryotlarda m. RNA sentezi (transkripsiyon) Splicing; Pre-m. RNA olgun m. RNA
GENETİK ŞİFRE
Genetik Şifre (Kod) Organizmalardaki proteinlerin birincil yapılarının (amino asit dizilerinin) DNA molekülündeki genlerin nükleotid dizisi tarafından tayin edilmesi ile ilgili şifre = DNA’nın nükleotid dizisi ile proteinlerin amino asit dizisi arasındaki ilişki. Genetik Şifrenin Özellikleri DNA (ve m. RNA) da proteinlerde 4 çeşit nükleotid 20 çeşit amino asit bir nükleotid bir amino asit iki nükleotid bir amino asit üç nükleotid bir amino asit (41 = 4 ) XXX (42 = 16) XXX (43 = 64) !!!!! Bir amino asidi belirleyen üç nükleotidlik (bazlık) dizi = kodon
Genetik Şifre (kodonlar RNA üzerinde ve 5’ 3’ yönünde yazılı. Buna göre DNA’daki tamamlayıcı kodonlar ters yönde (3 5 ), t. RNA antikodonları da m. RNA’dakinin tamamlayıcısı ve onunla ters yönde).
Genetik şifrenin yapısı ile ilgili önemli bir özellik: şifrenin okunma çerçevesinin her gende sabit bir başlangıç noktasından başlaması ve kodonların dizilişinin üste gelmemesi !!!!! Bu özellik tek çeşit ürünü şifreleyen genlerdeki kodonlar için geçerli; birden fazla genin üste bulunduğu bölgelerde genlerin okuma çerçeveleri farklı (ilk örnek, X 174 fajının genomundaki üste çakışan genler)
Kodonlar 61 kodon, amino asitleri şifreleyen kodonlar (anlamlı kodon) 3 kodon (UAA, UAG, UGA) bir amino asidi şifrelemeyen, protein sentezinde bitim (sonlandırıcı) işareti görevi yapan kodonlar [anlamsız kodon (bitim kodonu)]. Protein sentezini başlatıcı kodon genellikle AUG (metionini şifreleyen anlamlı kodon). Genetik şifrenin özelliği: yapısal benzerlik gösteren amino asitlerin genellikle benzer kodonlar tarafından tayin edilmesi. Örneğin, aromatik amino asit olan fenilalanin (UUU, UUC), tirozin (UAU, UAC), triptofan (UGG) kodonlarının urasil ile başlaması
Genetik şifredeki bozulma (dejenerasyon): aynı anlamı veren birden fazla kodonun bulunması. bir proteinin amino asit dizisinden giderek onu şifreleyen nükleotid dizisinin bilinmesi mümkün değil. Bozulma rastgele değil !!! Aynı amino asidi belirleyen kodonların genellikle 3. bazları farklı protein sentezindeki olası hataların en aza indirgenmesi !? !?
Genetik şifredeki bozulma
Kodonlarda bozulmanın özelliği farklı organizmaların, proteinlerindeki amino asit oranlarında önemli değişmeler olmaksızın, DNA’larındaki AT/GC oranında büyük farklılıkların olabilmesi. Genetik şifredeki bozulmanın nedeni: Salınım (titreşim, “wobble”) Hipotezi. Protein sentezi sırasında bir t. RNA molekülünün m. RNA ile ilişkisinin, kodonun 5’ ucu ile antikodonun 3’ ucu arasında başlaması; ilk iki pozisyonda doğru baz çiftleri oluştuğunda, üçüncü pozisyondaki hatalı baz eşleşmesinin önemli olmaması. Bir amino asidi taşıyan t. RNA’nın bazen aynı amino aside özgü birden fazla kodonu tanıyabilmesi.
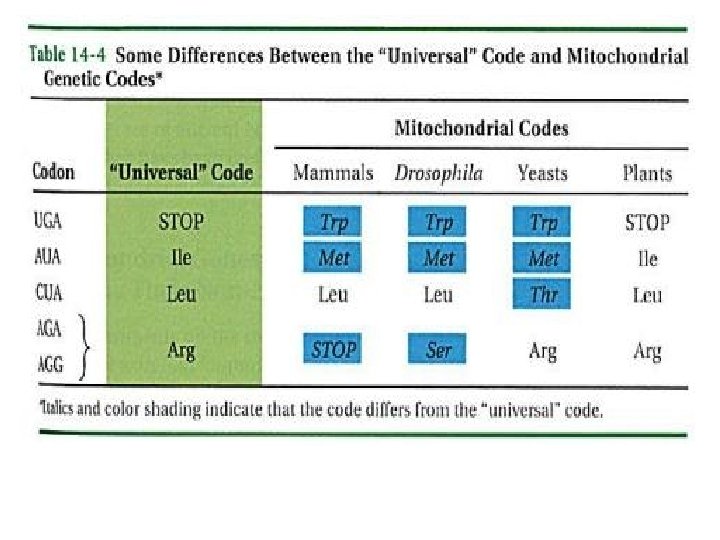
Genetik Şifrenin Evrenselliği Tüm organizmalarda genetik şifrenin genel biçimi ve işlemesi temelde aynı !!! Genetik şifrenin evrim süresi boyunca değişmeden kaldığının işareti. Şifrenin evrenselliğine ters düşen bazı farklılıklar: ▪Bakteriler ve ökaryotlarda AUG başlatma kodonunun anlamında küçük farklılık (bakterilerde formil metionin, ökaryotlarda metionin) ▪ Ökaryotlarda UGA’nın anlamsızlığı henüz gösterilmemiş ▪ Mitokondrilerde kullanılan şifrede temel genetik şifreye göre belirgin farklılıklar !!! ( memeli mitokondrilerinde UGA triptofan kodonu, AGA ve AGG bitim kodonları, AUG ve AUA metionin kodonları )
Referanslar • http: //www. microbiologyprocedure. com/genetics/chemical-nature-ofgenetic-materials/taylors-expreriment-on-seedlings-of-vicia-faba. htm • http: //www. emc. maricopa. edu/faculty/farabee/BIOBK/Bio. Book. DNAMOL GEN. html#DNA%20 Replication • www. istanbul. edu. tr/fen/mbg/notlar/