DNA TAMR Canllardaki genetik eitlilik bireylere evrim iin

DNA TAMİRİ

• Canlılardaki genetik çeşitlilik bireylere evrim için hammadde sağlar. • Genetik çeşitlilik nasıl oluşur? – Mutasyonlar – Rekombinasyonlar

MUTASYONLAR Mutasyon, genetik materialde meydana gelen kalıtılabilir ve tayin edilebilir değişikliklerdir. • Mutasyonlar evrimsel değişimin kaynaklarıdır. • Kendiliğinden veya mutajenler sonucu oluşur. • Genetik çeşitliliğin temelini oluştururlar. • Organizmalarda değişimler (mutasyonlar) yeni alleller ortaya çıkarır. • Mutasyon sonucu ortaya çıkan yeni allel 2. düzey çeşitlilik için hammadde olur çünkü

MUTASYON • Mutasyonlar şu özellikleri gösterirler: • 1. Kalıtsal maddedeki nukleotitlerin çeşit, sayı veya sırasında devamlı olan değişikliklerdir yani kalıtsaldır. • 2. Daha önceden şifrelenmemiş ya da programlanmamış değişikliklerdir. • 3. Oldukça ender meydana gelen değişikliklerdir. Çünkü kalıtsal madde şifrelenmemiş ender değişimlerin oluşumuna karşı çift sarmal yapısı ve proteinlere bağlanması ile kendini koruma eğilimdedir. • 4. Replikayonun doğruluğu ve onarım sisteminin etkinliği ile hata oluşumu en aza indirilmektedir.

Hücreler hasarlı DNA’yı onaracak biçimde evrimleşmiştir. Hücrelerde çeşitli onarım sistemleri vardır ve bunların çoğu da DNA’nın komplementer olma özelliğine güvenerek onarımı gerçekleştirir. Yani bir zincir kalıp olarak kullanır ve diğerini onarır.

MUTASYONLAR Mutasyonlar etkiledikleri dokulara göre de iki gruba ayrılırlar: - Somatik mutasyonlar - Germline (eşeysel) mutasyonlar

SOMATİK MUTASYON Somatik dokularda meydana gelen mutasyonlardır. Mutasyonlar mitoz bölünmesi sırasında DNA replikasyonu aşamasında meydana gelir. Sonradan ortaya çıkıp, kendi serisindeki hücreleri etkiler. Sonraki nesillere aktarılmaz. Birçok kanserin nedenidir. Germline mutasyonlar Eşey hücrelerinde meydana gelen mutasyonlardır. n Eşey dokularının olgunlaşması sırasında mitozda ve mayozda meydana gelir. n Meydana gelen mutasyon sonraki nesillere aktarılır. n
 İntronlarda Ekspresyon kontrol")
MUTASYONLAR DNA’DA NERELERDE MEYDANA GELİR? 1. Gen Bölgesinde Kodlanan dizilerde (eksonlarda) İntronlarda Ekspresyon kontrol bölgelerinde Buralarda meydana gelen mutasyonlar son ürünün yapısını veya ekspresyon düzeyini etkiler 2. Genlerarası Dizilerde Buralarda meydana gelen mutasyonlar nötr olarak bilinse de, günümüzde bazı önemli etkilerin veya biilmediğimiz bazı kontrol mekanizmaların etkilendiği söylenebilir.

MUTASYONLAR Mutasyonlar genel olarak iki gruba ayrılırlar: - Tek gen mutasyonları - Kromozomal mutasyonlar

MUTASYON • 1. Gen Mutasyonları: Genlerin yerinde değişme olmaksızın Mutasyonları: yapılarında meydana gelen değişmelerdir. Mutasyon = Gen Mutasyonu • Moleküler düzeydeki tanımıyla gen mutasyonu; genin yapısını oluşturan nukleotidlerin sayısında, oranında, sıralamasında meydana gelen değişimlerdir.

GEN MUTASYONLARI Genin yapısındaki değişmeyle taşıdığı şifre değişeceği için o gene özgü ürün de değişime uğrar § Bu değişme bazen fenotipte gözlenmeyecek kadar önemsiz olabilir. § Bazen protein biraz değişebilir § Ya da genin fonksiyonu mutasyonla tamamen kaybolabilir

GEN MUTASYONLARI Gen mutasyonları, bir gende meydana gelen genellikle küçük değişikliklerdir. Gen mutasyonları kendi içinde iki gruba ayrılırlar: -Nokta mutasyonları -Çerçeve mutasyonları

NOKTA MUTASYONLARI Nokta mutasyonları, DNA dizisinin herhangi bir bazında meydana gelen değişikliklerdir. Bu değişiklik tek bir bazda meydana gelmektedir. Nokta mutasyonu değişen bazın dönüştüğü baz türüne göre iki gruba ayrılmaktadır: Transisyon › Purin bazın purine A --> G veya G --> A › Pirimidin bazın pirimidine C --> T veya T --> C Transversyon › Purin bazların pirimidine A veya G --> T veya C › Pirimidin bazların purine T veya C --> A veya G

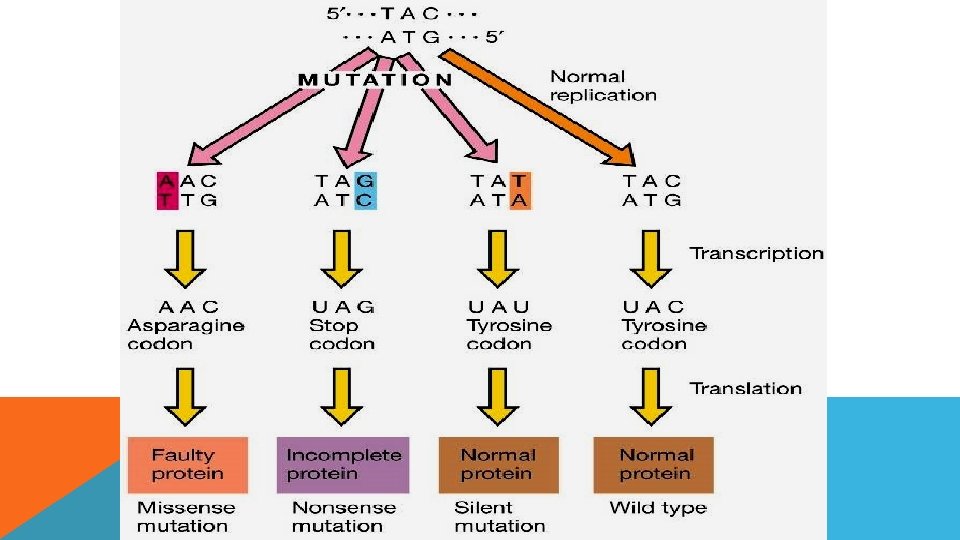
NOKTA MUTASYONLARI Nokta mutasyonları sonucu, bir genin nükleotit dizisindeki bazlarda meydana gelen değişiklikler sonucunda genetik kodonlardan birinde değişiklik olur. Nokta mutasyonları kodon değişimleri sonucuna göre üç gruba ayrılırlar: - Eş anlamlı mutasyonlar (Sessiz mutasyon) - Yalnış anlamlı mutasyonlar - Anlamsız mutasyonlar

NOKTA MUTASYONLARI Eş anlamlı mutasyonlar sonucu kodonda değişiklik olsa da kodlanan amino asit değişmez. Ör, UAC > UAU >Tyr Yalnış anlamlı mutasyonlar sonucu kodonda meydana gelen değişiklik sonucu kodlanan aa değişir. Ör, UAC > Tyr, AAC>Asn Anlamsız mutasyon sonucu kodonda meydana gelen değişiklik stop kodonu oluşturur. Ör, UAC > Tyr, UAG > Stop

REVERS MUTASYON 1 ATG met 4 GGA gly 7 GCT ala ATG met GGA gly GCT ala 10 CTA leu 13 16 19 TTA ACC TAA leu thr stop 1. mutasyon CTA TTT ACC TAA leu phe thr stop 2. mutasyon CTA TTA ACC TAA leu thr stop veya 2. mutasyon CTA CTT ACC TAA leu thr stop

NOKTA MUTASYONLAR Genin işlevi kaybetme olasılığı yeni bir genin işlev kazanmasından daha olasıdır. Ancak işlev kazanma olasılığı da vardır

ÇERÇEVE MUTASYONLARI Çerçeve mutasyonları, gen dizisindeki baz sayısında artma veya azalma sonucu ortaya çıkan mutasyonlardır. Çerçeve mutsyonları kendi içinde üç gruba ayrılır: - İnserisyon - Delesyon - Kayma

ÇERÇEVE MUTASYONLARI İnsersiyon, gen dizisine üç ve katları şeklinde nükleotit eklenmesidir. Delesyon, gen dizisinden üç ve katları şeklinde nükleotit çıkarılmasıdır. Kayma mutasyonları, üçe bölünmeyen sayılar kadar nükleotitin gen dizisine eklenmesi veya çıkarılmasıdır.
")
Orjinal Dizi AUG CAC GGU AAC GCG CCG GUU AAA UGG Met-His-Gly-Ser-Asn-Ala-Pro-Val-Lys-Trp İnsersiyon (Duplikasyon) Delesyon AUG CAC GCG GGU AAC GCG CCG GUU AAA UGG Met-His-Ala-Gly-Ser-Asn-Ala-Pro-Val-Lys. Trp AUG CAC GGU AGU GCG CCG GUU AAA UGG AAC Asn Met-His-Gly-Ser-Ala-Pro-Val-Lys-Trp AUG CAC GGU AAC AGC GCC GGU UAA AUG G Kayma (frameshift) Met-His-Gly-Ser-Asn-Ser-Ala-Val-Stop

Mutasyon tipi Örnek Normal İKİ KAŞ İKİ GÖZ BİR YÜZ ÇİZ Yalnış Anlamlı İKT KAŞ İKİ GÖZ BİR YÜZ ÇİZ Anlamsız İKİ KAŞ İKİ Delesyon İKİ GÖZ BİR YÜZ ÇİZ Insersiyon İKİ KAŞ İKİ GÖZ GÖL BİR YÜZ ÇİZ Duplikasyon İKİ KAŞ İKİ GÖZ BİR YÜZ ÇİZ Kayma İKİ KAŞ İKD İGÖ ZBİ RYÜ ZÇİ Z

FARKLI MUTASYONLAR – AYNI HASTALIK LDL reseptöründeki mutasyonar reseptörün işlevini bozar, kan kolestrolünün yüklesmesine ve erken kalp hastalığına neden olur (Ailesel hiperkolestrolemi)

KROMOZOMAL MUTASYONLAR • 2. Kromozom Yapı veya Sayısını Değiştiren Mutasyonlar • a- Kromozom Sayı Değişimleri (Genom Mutasyonları) • Kromozom takımları sayısında tam katlar halinde artma (poliploidi) veya azalmalarla (monoploidi) takımdaki kromozomlardan bazılarının sayısındaki artma veya azalmaları (anöploidi) kapsar. • b- Kromozom Yapı Değişmeleri (Kromozom Mutasyonları) • Kromozomlarda kırılmalara sonucu oluşan parça kayıpları (delesyon) delesyon veya artışlarını (duplikasyon), duplikasyon parça yerleşim düzenlerindeki değişimleri (inversiyon) inversiyon ve kromozomlar arası parça değiş tokuşlarını (translokasyon) translokasyon kapsar. Bu tip mutasyonlar kromozomların sayısını veya kromozomlardaki geniş bölgeleri ilgilendiren büyük değişimlerdir. Bu tür değişimlerle genlerin ya sayısı ya da yerleşim düzenleri değişir bunun sonucunda da bireyin fenotipinde kalıcı değişimler ortaya çıkar
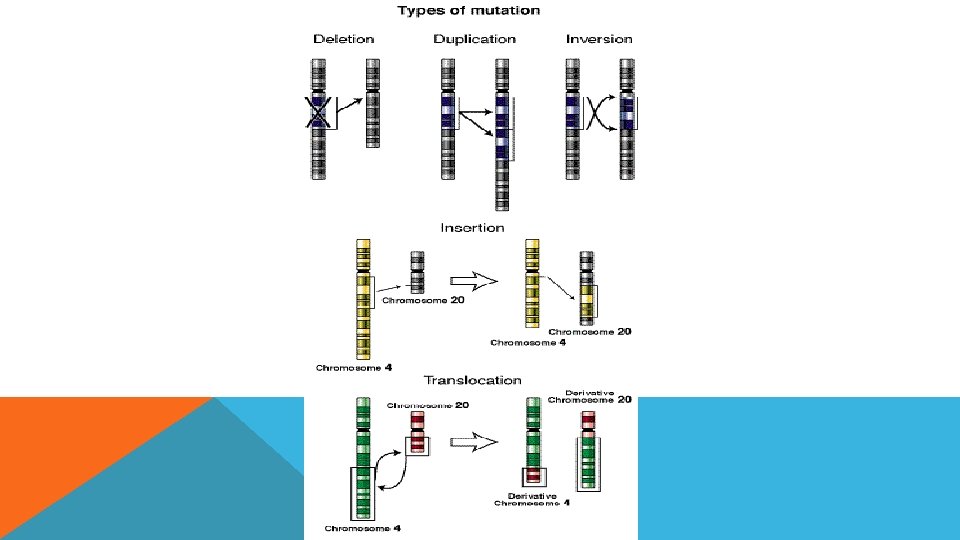

YAPISAL MUTASYONLAR Yapısal mutasyonlar sonucu kromozom yapısında değişiklikler meydana gelir. Yapısal mutasyonlar kendi içinde beş gruba ayrılır: - Delesyon - Duplikasyon - İnsersiyon - İnversiyon - Translokasyon

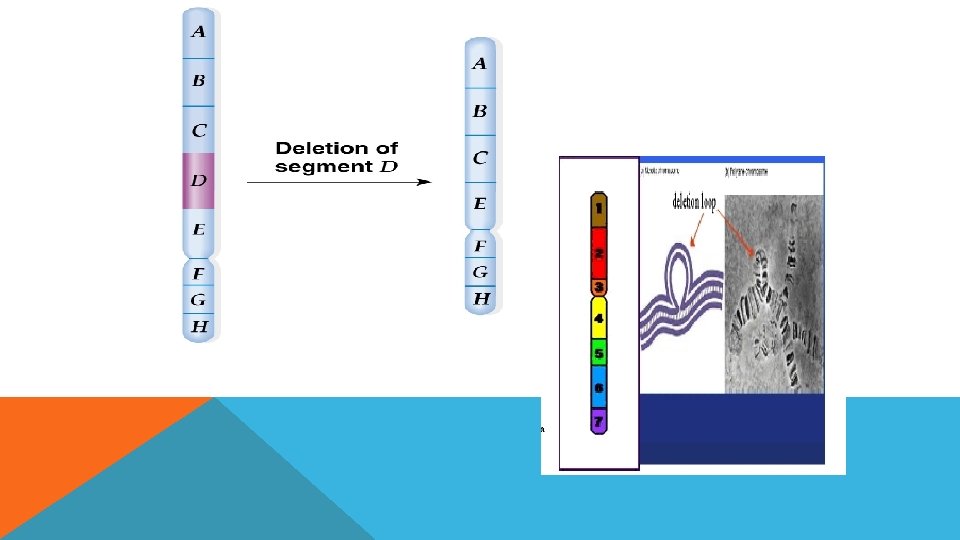
DELESYON Delesyon kromozomdan parça kopmasıdır. Kromozomun herhangi bir bölgesinin veya kaollardan herhangi birinin kopmasıyla meydana gelir.

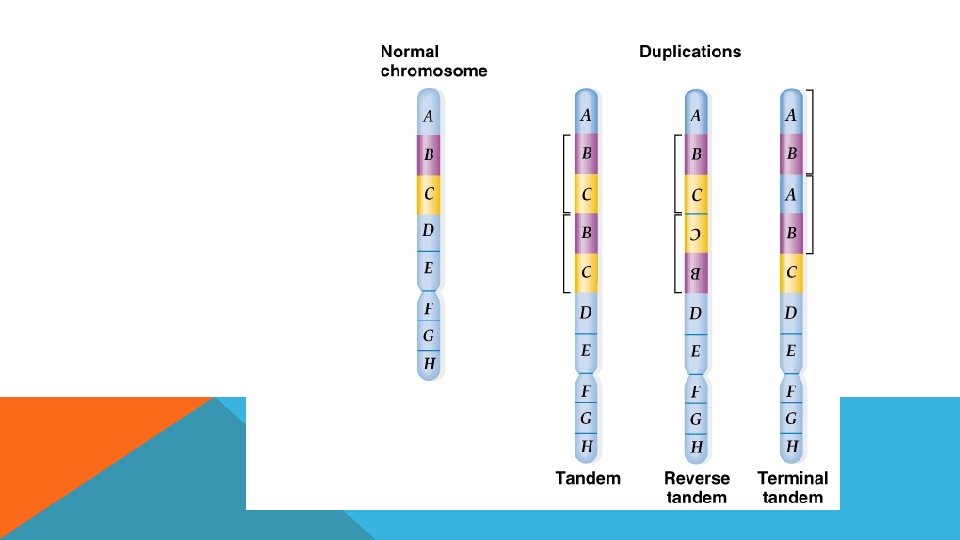
DUPLİKASYON Kromozomu herhangi bir bölgesinin fazladan bir kopyasının genomda bulunmasıdır.

İNSERSİYON Kromozomun herhangi bir bölgesine frklı bir genom parçasının entegre olmasıdır. Insertion mutations 5' 3' 3' 5'
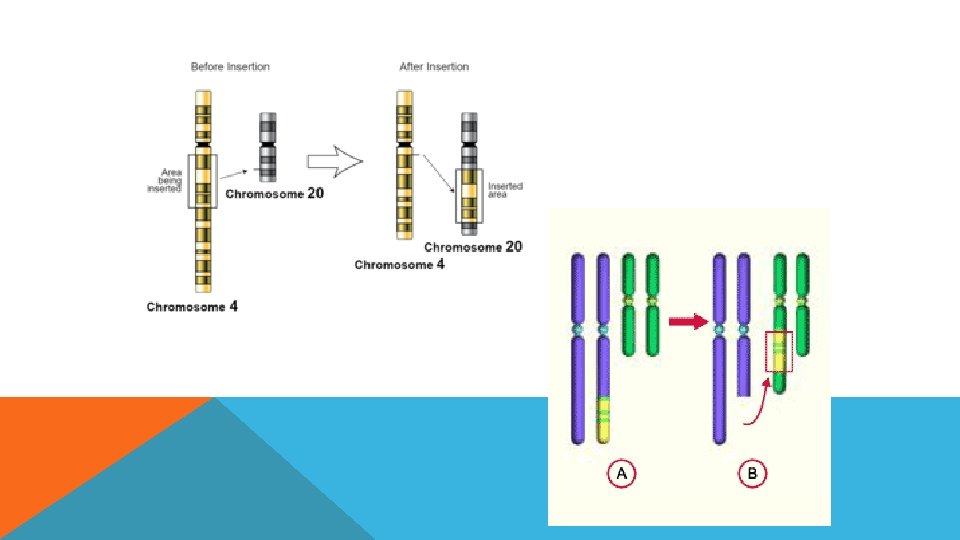

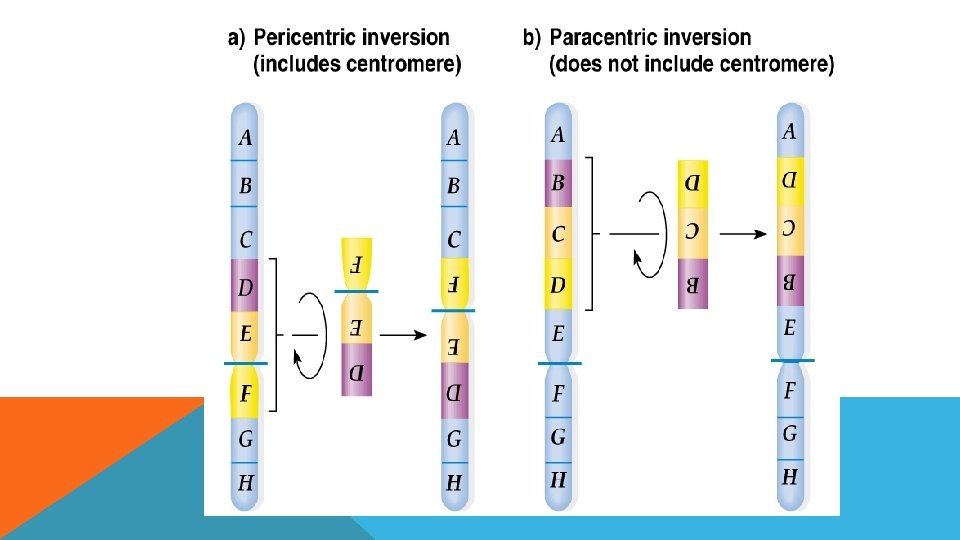
İNVERSİYON Kromozomun herhangi bir bölgesinin koparak kendi ekseninde dönüp tekrar kromozoma entegre olmasıdır.

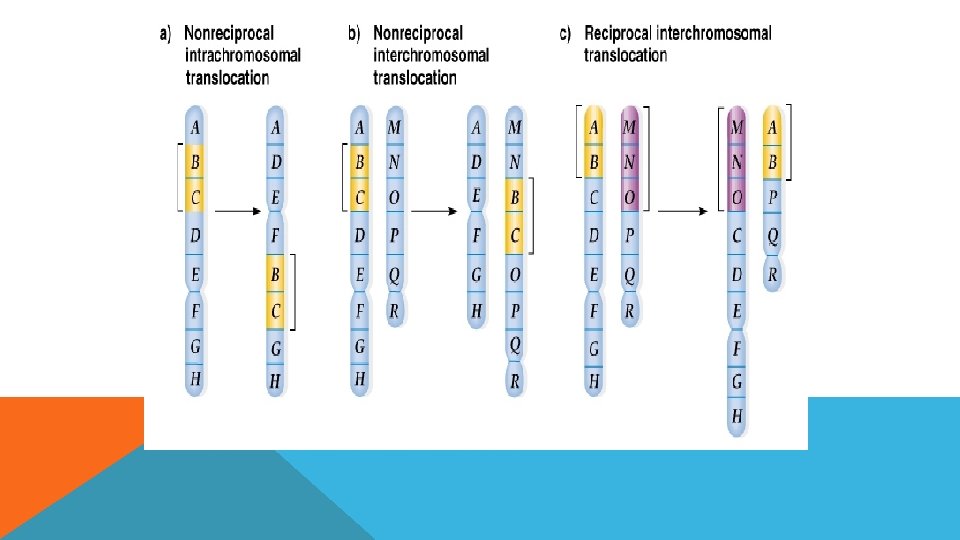
TRANSLOKASYON Herhangi iki farklı kromozom parçasının kendi aralarında yer değiştirmeleridir.

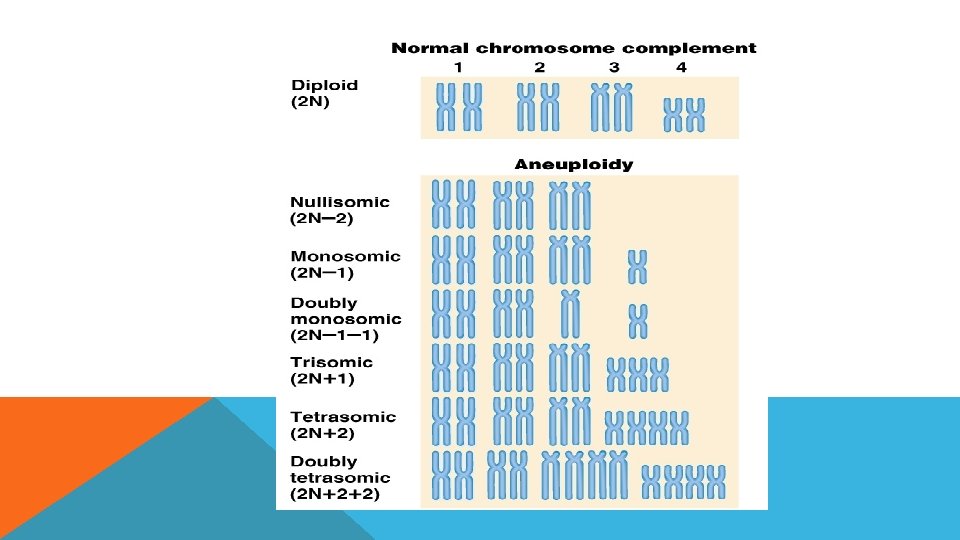
SAYISAL MUTASYONLAR Sayısal kromozomal mutasyonlar sonucu canlı genomunda bulunan kromozomların sayısında değişiklikler meydana gelir. Sayısla mutasyonlar kendi içinde iki geuba ayrılır: - Aneupolidi - Polipolidi

ANEUPLOİDİ Aneuploidi, genomda bulunan kromozomlardan biri veya bir kaçının artışı veya azalışıdır.
 : Kromozom anomalilerini meydana getiren mekanizmalardan en")
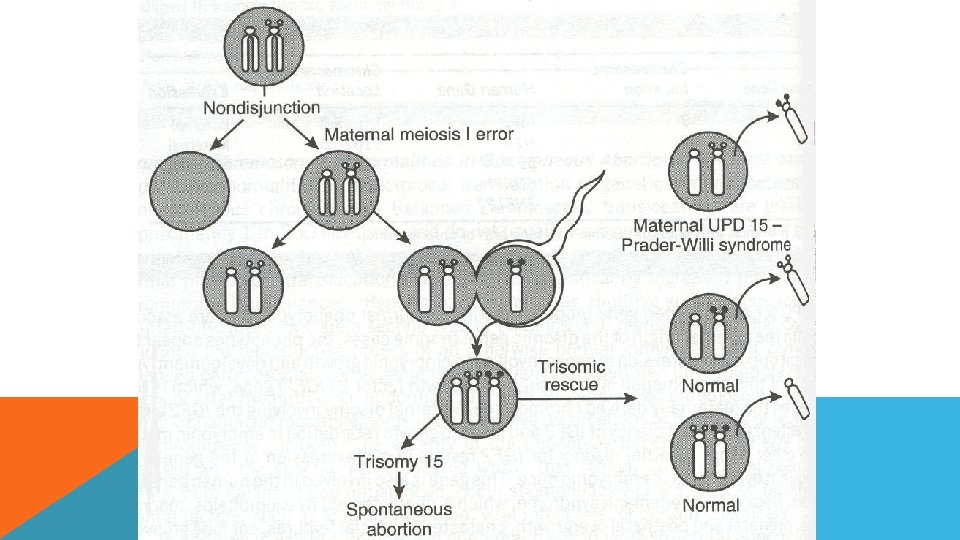
ANEUPLOİDY OLUŞ NEDENLERİ 1. Kromozom ayrılmaması (nondisjunction) : Kromozom anomalilerini meydana getiren mekanizmalardan en önemlisi mayozda gametlere az, ya da çok sayıda kromozom gitmesidir. Bu durum ya mayoz ya da mitoz bölünmede olabilir. Meydana gelen bu olayın sonunda kromozom anöploidisi oluşmaktadır. Nedeni ileri anne-baba yaşı, viral enfeksiyonlar, kimyasal ajanlar ve radyasyondur
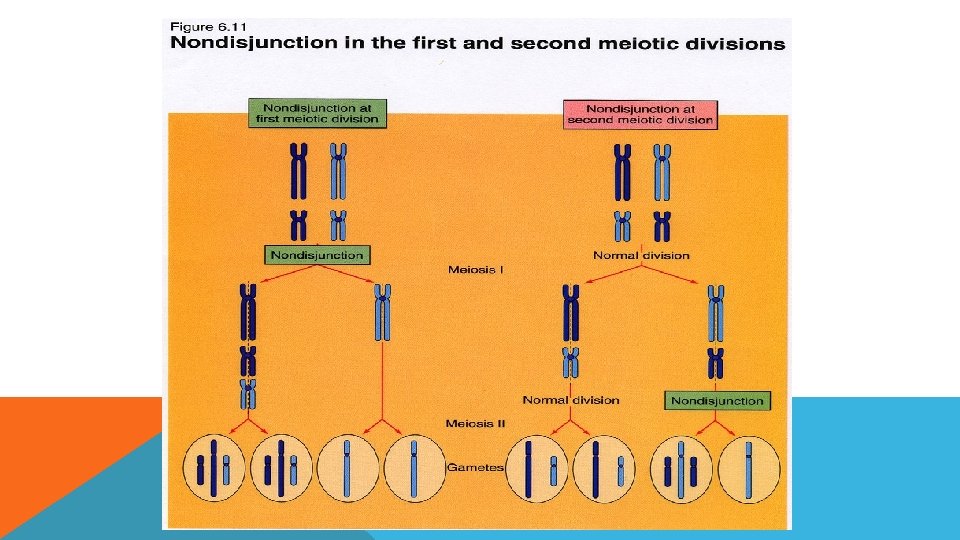
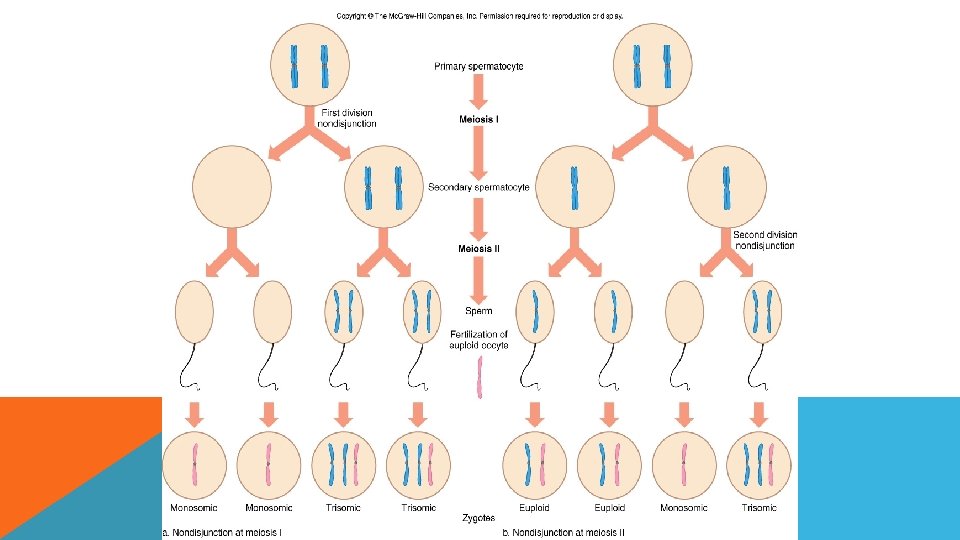
 : Normal olarak")
ANEUPLOİDY OLUŞ NEDENLERİ 2. Kromozomların anafazda geri kalması (anaphase lagging ) : Normal olarak uzunlamasına bölünerek kutuplara çekilmekte olan kromozomlardan biri anafazda geri kalır. Hareket etmekte geç kalan bu kromozom ya özdeşinin bulunduğu kromozom grubuna katılır yada bölünme sırasında ortadan kaybolur.
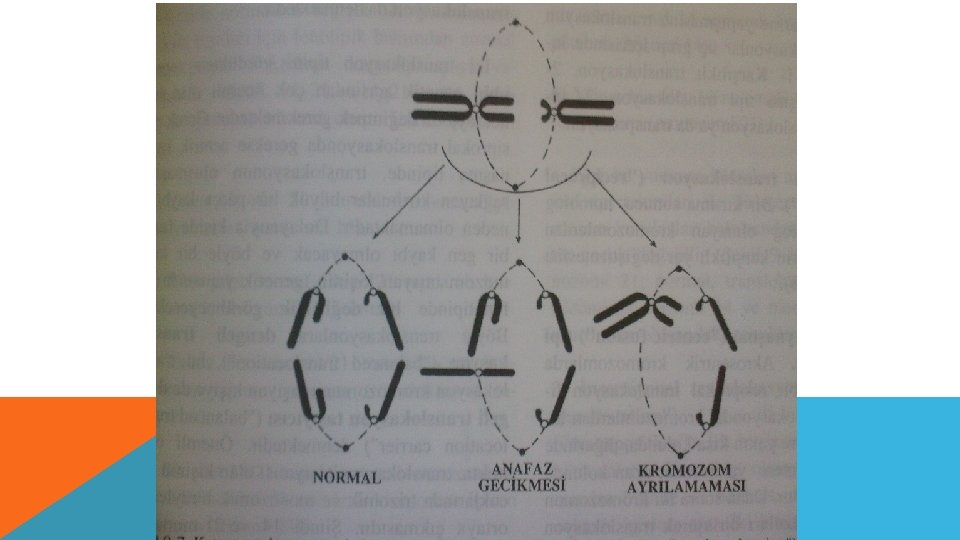
, normal karyotipte, fakat herhangi bir kromozomunun her iki")
UNİPARENTAL DİZOMİ UPD (Uni. Parental Dizomi), normal karyotipte, fakat herhangi bir kromozomunun her iki kopyasının da ayn ebeveyinden gelmesi olayıdır. UPD çeşitleri: - Heterodizomi - İzodizomi
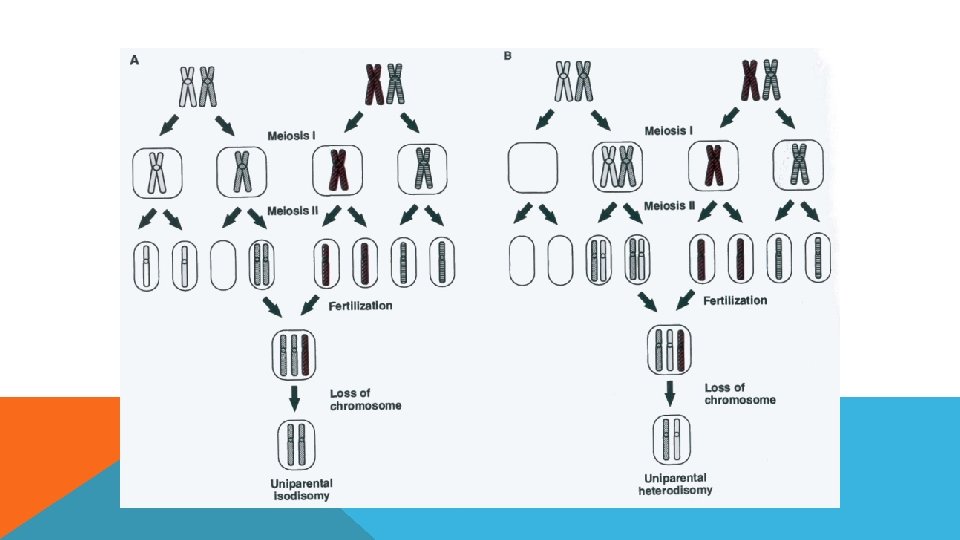
, canlının kromozom setinde artış veya azalıştır.")
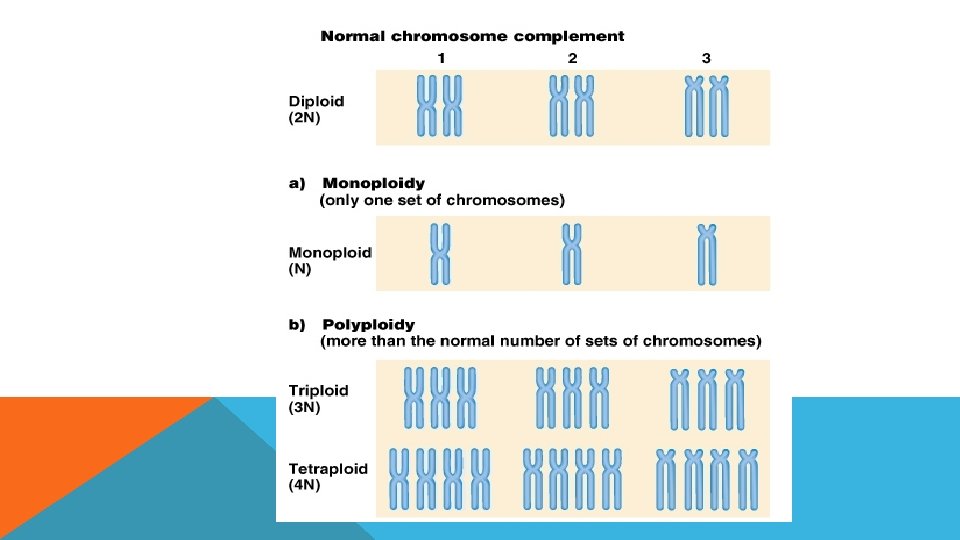
POLİPLOİDİ Poliploidi (öploidi), canlının kromozom setinde artış veya azalıştır.

TÜM MUTASYONLAR ZARARLIMIDIR? Mutasyonlar her zaman zararlı değildir. Canlı için yararlı mutasyonlar da vardır. § Örneğin, hücre yüzey proteininde (CCR 5) meydana gelen mutasyon HIV enfeksiyonuna direnç kazandırır. Mutasyonlar, ayrıca evrimin başlıca nedenidir.

MUTASYONLAR Mutasyonlar oluşum durumlarına göre beşe ayrılırlar: 1 - Spontan – sporadik 2 - Kromozomlararası yeniden düzenlemelerle meydana (rearrangement) gelen 3 - Mitotik ve mayotik hatalar (nondisjunction) sonucu oluşan 4 - Kalıtsal (ailesel) Mutasyonlar 5 - İndüklenmiş Mutasyonlar

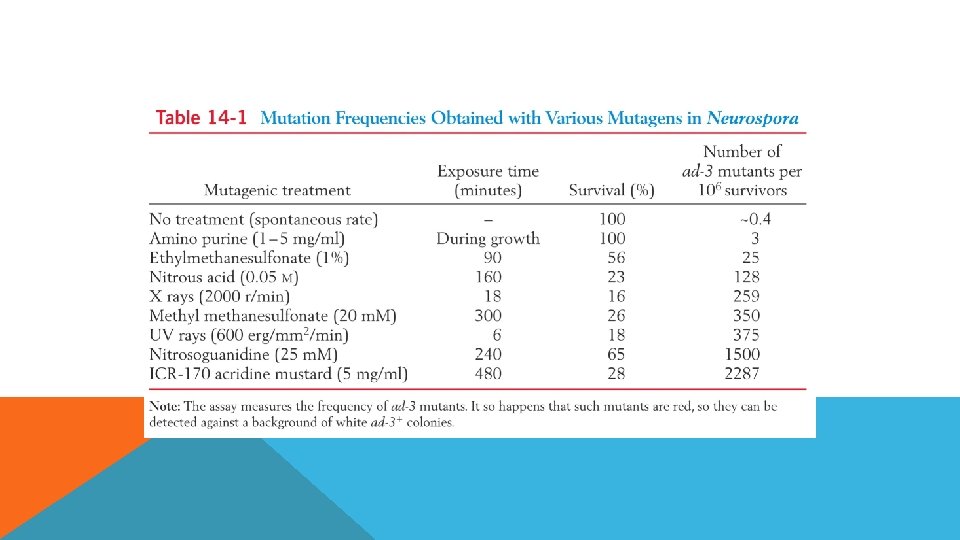
MUTASYONLAR Spontan mutasyonlar, canlının veya hücrenin normal yaşamı boyunca DNA polimerazın hata payı, nükleotitleri totomerizasyonu, oksidatif deaminasyon gibi nedenlerle meydana gelen mutasyonlardır. Mutajenlerin etkisi yoktur. Spontan mutasyon hızı ortalama 10 -6 düzeyindedir. İndüklenmiş mutasyonlar, mutajen olarak isimlendirilen fiziksel veya kimyasal ajanların etkisiyle oluşan mutasyonlardır

MUTAJENLER Spontan mutasyon frekansı oldukça düşüktür bir hücrede yaklaşık 105 -108 hücrede bir görülür. Dolayısıyla daha fazla mutasyon isteniyorsa mutlak indüklemek gerekir. Mutajenlere maruz bırakarak mutasyon üretmeye mutagenez denir ve bu organizmaya da mutagenize organizma denir.

MUTAJENLER En sık kullanılan mutajenler Yüksek enerjili radyasyon Çeşitli kimyasallar § Mutajenin dozu nekadar yüksekse mutasyon freakansı da okadar yüksek olur

NOKTA MUTASYONLARIN İNDÜKLENMESİ • Her mutajen farklı bir mutasyon özgüllüğü yada önceliğine sahiptir • DNA üzerindeki hedeflenen mutasyon bölgelerine yada mutasyona yatkınlık gösteren bölgelere sıcak nokta denir (hot spot) • Mutajenler üç farklı mekanizmayla etkilerini gösterirler • Baz başka bazla değişmesi • Bazın hasar görmesi

SPONTAN MUTASYON MEKANİZMALARI Replikasyon hataları Spontan lezyonlar Transpoze elementlerin insersiyonu § Spontan mutasyonlar nadir görülür ve onların altındaki nedeni belirlemekde zordur

SPONTAN LEZYONLAR Doğal olarak DNA’nın hasara uğramasına spontan lezyonlar denir ve bu hasarlar mutasyona neden olurlar En sık rastlananları § Depurinasyon § deaminasyon
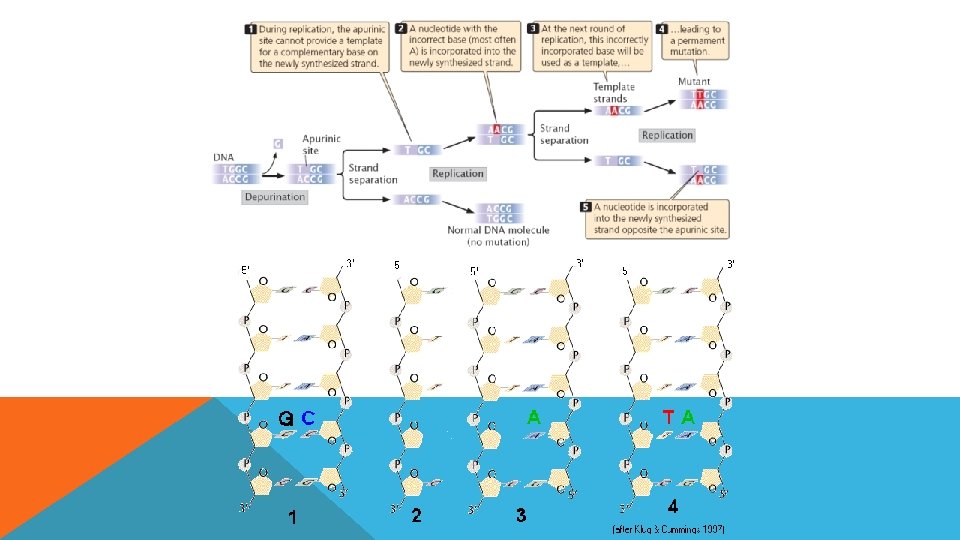

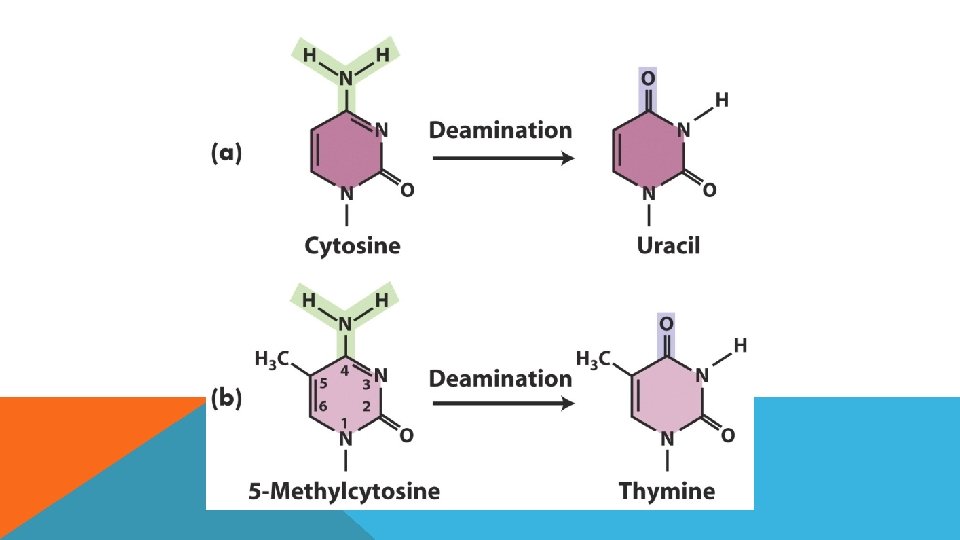
DEAMİNASYON • Sitozin deaminasyonu urasil üretir • Düzeltilmedikçe replikasyon sırasında urasil adenin ile eşleşir ve GC çifti AT ile değişir. • 5 -metil sitozin deaminasyonu da görülür çünkü pro ve ökaryotlarda bazı bazlar metillenirler. • 5 -metil. C deaminasyonu timin üretir (5 metilurasil) dolayısıyla C-T transisyonu deaminasyonla üretilir. • Adenin deaminasyon ile Hipoksantin’e dönüşür ve Sitozin ile eşleşir.

OKSİDATİF DEAMİNASYON

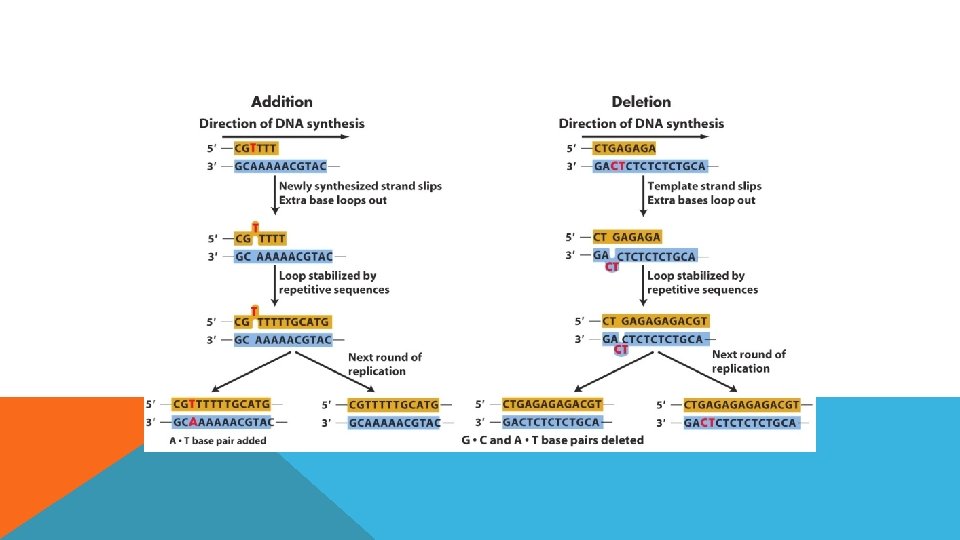
DNA REPLİKASYON HATALARI Baz değişimi § Hiçbir kimyasal reaksiyon mükemmel değildir dolayısıyla DNA sentezi sırasında da yanlış nt çiftleri oluşturulabilir. Baz insersiyonu ve delesyonu Replikasyon sırasında indel mutasyonlarda görülür eğer üçlü nt biçiminde değilse frame kaymasına neden olur Bu eklenti yada çıkarılmalar replikasyon kayması olarak da bilinir. Genellikle art arda dizilmiş aynı nt lerin olduğu bölgelerde bu kaymalar meydana gelir.

TOTOMERİZASYON A T keto TG enol C G A T

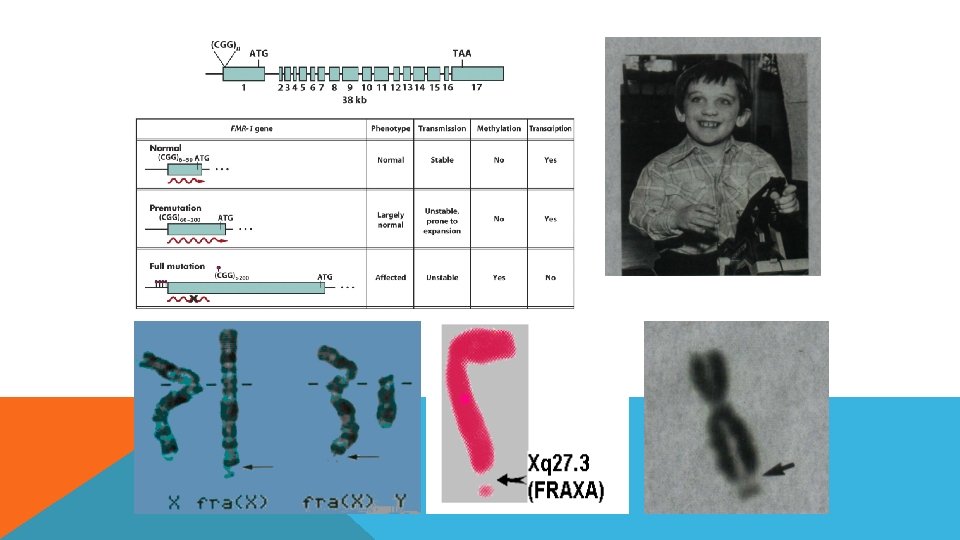
İNSANDA SPONTAN MUTASYONLAR-TRİNÜKLEOTİD TEKRARLARI • İnsanda görülen birçok hastalığın etkeni karmaşık mutasyonlardan kaynaklanır. Bunlardan en önemlilerinden biri kısa tekrarların duplikasyonlarıdır. • Birçok kalıtsal hastalıkdan sorumlu genetik defektin altında yatan mekanizma 3 lü nt dizilerinin tekrarıdır (trinucletid repeat) • Fragil X sendromu buna örnektir • 1/1500 erkek ve 1/2500 dişide görülür sitogenetik olarak kromozom incelenmesinde x kromozomunda kırılganlıklar görülebilir. • Frajil X FMR 1 genindeki CGG tekrarlarındaki değişim sonucu ortaya çıkar bu gen transkribe olur ama translase olamaz. • Tekrar sayısı hastalık fenotipiyle nasıl doğru orantılı biçimde seyreder? • İnsanlar doğal olarak FMR 1 genindeki CGG tekrarlarında çeşitlilikler gösterir ve bu tekrarların sayısı 6 -54 arasında değişir. Bazan etkilenmeyen ebeveynlerden hatta büyük baba/anneler kendinden sonraki kuşaklarda etkilenmiş çocuk sahibi olurlar. Bu hasta bireylerde tekrar sayıları 200 -1300 e ulaşır. Etkilenmemiş ebevynlerde de bazan sayı 50200 arasında değişir. Dolayısıyla bu etkilenmemiş ebevynler premutasyonu taşımaktadırlar. Premutantlardaki Bu tekrarlar hastalık fenotipinin ortaya çıkması için yeterli değildir ancak bu kişiler oldukça yatkın kişilerdir.

TEKRAR MEKANİZMASI DNA sentezi sırasında eşleşme kaymasından kaynaklanır. İnsanda 50 tekrardan sonra DNA Pol enziminin doğru sentez yapmadığı ileri sürülmektedir. Yani 50 tekrar DNA pol için eşik değeridir. Bu noktadan sonra mutasyon sıklığı büyük ölçüde artar.

MUTAJENLER I- Fiziksel Mutajenler a- Isı b-p. H c- Işınlar 1 -İyonize ışınlar (X ve gamma) 2 -Non-iyonize (UV, 260 nm dalga boyu ışınlar) 3 -Mor ötesi ışınlar

Elektomanyetik Radiasyon Gamma-ışınları, Xışınlsrı - İonize radyasyon UV ışını - DNA zincirini kırar - Primidin dimerine neden oluyor

Nonionize Radiasyon n. Non-ionize radiasyonun en etkili dalga boyu 260 nm’ye kadar olan UV dalgalarıdır. UV DNA zincirinde Primidin (Timin) dimerilerinin oluşumuna neden olur. n. Replikasyon sırasında DNA Polimeraz Timin dimerileri bölgesinde duraklar ve yanlış nükleotid ekler.

Elektomanyetik Radiasyon ?

MUTAJENLER II-Kimyasal Mutajenler a- Baz Analogları (5 -Bromodeoksiuridin-Brd. U, 6 -thioguanin, 2 aminopürinler en yaygınları) b- Deaminasyon yapan ajanlar: DNA yapısında amino gruplarının kaybına neden olan ajanlar (Nitröz asidi, hidroksil aminler) c- Alkilleyici ajanlar: DNA yapısına alkil grubu takan ajanlar ( Kükürt, Nitrojen mustard, Etilenoksitler) d- İnterkalasyon yapan ajanlar : Acridinlerin hepsi bu özelliktedir acrilflavin ve acridin orange (Proflawin, e- Demetilasyon yapan ajanlar: DNA’nın hipo yada demetilasyonuna neden olan ajanlar (5 azacytidine, 5 aza-2 -deoxycytidine) f- Çeşitli insersiyonlara neden olan ajanlar ( Bu grup ajanlar DNA replikasyonu esnasındasüresince pürin ve primidin bazları yerine DNA yapısına katılan çoğunlukla frame-shift mutasyonlara neden olan ajanlardır. (ethidium bromide-Et. Br).

Baz Analogları n. Normal bazlara benzer ve replikasyon sırasında DNA’nın yapısına girer n 5 -bromouracsil – Keto formunda bulunur, ama sık enol formuna dönüşebilir –Keto formunda Adeninle bağ oluşturur –Enol formunda Guaninle –Sonuçta AT’den GC’ye dönüşen trasisyon mutasyon oluşur

BAZ ANALOGLARI Baz analogu yapısı eşleşme Keto formu: -A Enol formu: -G -T -C normal Baz

Interşalasyon Ajanlar n. Bu mutajenler DNA çift zincirinin arasına girerek mutasyon yaprlar Örnek ajanlar; acridine orange, ethidium bromid n. Replikasyon sırasında bu ajanların girdiği bölgede nmikroinsersiyon/delesyon veya kayma mutasyonları gözlenir.
 • Radyasyon (UV,")
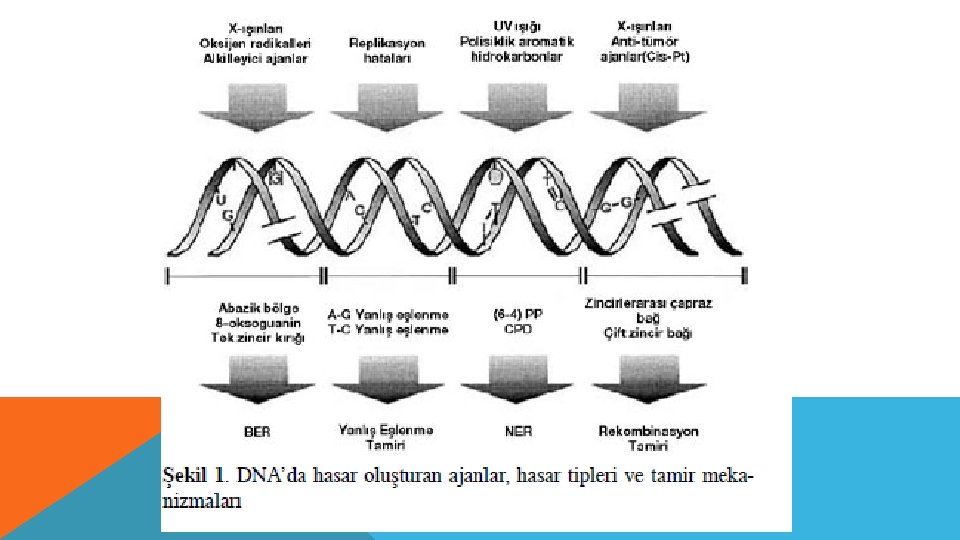
MUTAJEN-MUTASYON ÖRNEKLERİ Hasar Kaynağı: • sıcaklık • Metabolik Olaylar (srbest radikallar) • Radyasyon (UV, XRay) • Bileşenlerin etkisi (özellikle aromatik bileşenler) Hasar Şekli: • Nükleotit deaminasyonu • Nükletidlerin depurinasyonu • Baz oksidasyonu, DNA kırıkları • DNA zincirinde kırıklar

DNA TAMİRİ DNA tamiri için bilinen beş farklı tamir mekanizması vardır: Fotoreaktivasyon Yalnış eşleme onarımı Baz çıkarım onarımı Nükleotit çıkarım onarımı Rekombinasyonel onarım Çift Zincir Kırıklarının Tamiri

DNA HASARI VE TAMİRİ Hasarın Nedeni Hasarın Şekli Tamir Mekanizması Spontan modifikasyon, oksidasyon, alkilasyon, tek zincir kırıkları Tek zincirde tek baz hasarı Baz Ekzisyon Repair UV-ışın, zincir içi çapraz bağlanmalar, Tek zincirde birden çok baz hasarı Nukleotid Ekzisyon Repair X-ışını, zincirler arası çapraz bağlanma Her iki zinciri etkileyen hasar Serbest Uçların nonhomolog bağlanması (NHEJ) Homolog rekombinasyon (HR) Replikasyon hataları ve kaymalar Yanlış eşleşmeler, insersiyon ve delesyonlar Mismatch (yanlış eşleşme) eksizyon tamiri MER

DNA TAMİR MEKANİZMALARI İLE İLİŞKİLİ 100’ÜN ÜSTÜNDE GEN OLDUĞU TESBİT EDİLMİŞTİR. Tamir mekanizması Ana genler Yardımcı genler 1 0 Mismatch (yanlış eşleşme) eksizyon tamiri 22 4 Nukleotid Ekzisyon Repair 32 5 Baz Ekzisyon Repair 32 7 Homolog rekombinasyon 19 1 Direkt tamir Serbest Uçların non-homolog bağlanması Homolog rekombinasyon
 - Özellikle DNA yapısında en yaygın meydana gel “timin dimer” mutasyonunun")
FOTOREAKTİVASYON (Direkt tamir) - Özellikle DNA yapısında en yaygın meydana gel “timin dimer” mutasyonunun giderildiği tamir mekanizmasıdır. Görünür ışın (güneş ışını) aracılığıyla başarılır. _______________ A T G A C ıı ıı ııı T A C T G A A G ııı T = T Görünür ışın (güneş) C Mutant DNA zinciri(Timin dimeri) Fotoreaktivasyon (aktif DNA fotoliaz inaktif DNA fotoliaz) ________________ A T G A C A A G ıı ıı ııı T A C T G T T C Tamir edilmiş DNA zinciri fotoliaz (+görünür ışık)

Alkilasyon hasarının tamiri • Nitrozaminler ve N-metil-N 1 –nitrosoguanidin en önemli alkilleyici ajanlardır. Nukleotidlerdeki amino ve keto gruplarına metil (CH 3 – ) ya da etil (CH 3 – CH 2 ) gibi bir alkil grubu eklerler. • En önemli alkilasyon bölgesi, guaninin 6. karbon atomundaki oksijendir. Alkilasyon sonucunda oluşan O 6 –etilguanin (ya da O 6 -metilguanin), adeninin baz analoğu gibi davranarak timinle eşleşir. Bunun sonucunda hasarlı DNA replike olduğunda G: C baz çifti yerine A: T baz çifti geçer.

DİREK TAMİR
 • Özellikle replikasyon sonrası oluşan yalnış eşleşmeleri tamir")
YALNIŞ EŞLEŞME ONARIMI (MİSMATCH REPAİR MMR) • Özellikle replikasyon sonrası oluşan yalnış eşleşmeleri tamir eder • Prokaryot ve ökaryotlarda benzer mekanizmalar ve korunmuş protein yapısı söz konusudur. • Spontan mutasyon hızın 10 -1000 x katı düşürür. • İnsanlarda eksikliği kolon ve diğer kanser oluşumlarına yol açar

MMR Mekanizması
 DNA replikasyonunu bloke eden büyük lezyonların onarımında iş")
NUCLEOTİD ÇIKARIM ONARIMI (NUCLEOTİDE EXCİSİON REPAİR-NER) DNA replikasyonunu bloke eden büyük lezyonların onarımında iş yapar Yanlış eşleşme onarımına benzer E. coli’de, Uvr. ABCD enzim sistemiyle gerçekleştirilir Bazı proteinler ökaryotlarda da bulunmuştur İnsada bu onarımın buzukluğunda “Xeroderma pigmentosum” adlı hastalık gözlenir.
 Nukleotid ekzisyon tamiri E. Coli’de 4 protein ile gerçekleştirilir: Uvr.")
Nükleotid Eksizyon Tamiri (NER) Nukleotid ekzisyon tamiri E. Coli’de 4 protein ile gerçekleştirilir: Uvr. A, Uvr. B, Uvr. C ve Uvr. D. Uvr. A ve Uvr. B kompleksi, DNA’yı tarar. Uvr. A helikste yapısal bozuklukları belirlemekten sorumludur. Bir bozukluklukla karşılaştığında Uvr. A kompleksten çıkar ve Uvr. B lezyonun bulunduğu kısımda DNA’yı yumuşatır, açar ve ardından Uvr. C’yi çağırır. Uvr. C iki yarık oluşturur. Bir tanesi lezyonun 5’ kısmından 8 nukleotid uzaktadır Diğeri 3’ kısmında 4 ya da 5 nukleotid uzaktadır. Bu yarıklanma yaklaşık 12 -13 baz uzunluğunda tek sarmal DNA segmentini oluşturur. Bu segment, Uvr. D (DNA helikaz) ile uzaklaştırılır Oluşan boşluk, DNA Pol I ve ligaz ile doldurulur.

Tanıma ve bağlanma Kesme Çıkarım ve tamir
 desoksiribo fosfodiesteraz U-DNA glikozilaz Methyl-A 8 -Oxo-G Alk. A")
Baz çıkarım onarımı (değişik/modifiye bazlar) desoksiribo fosfodiesteraz U-DNA glikozilaz Methyl-A 8 -Oxo-G Alk. A Mut. M AP endonükleaz DNA pol I

Recombinasyon Onarım DNA pol. durmaz -Replikasyon gerçekleşir -Daha sonra onarım olur Lesion TT-dimer
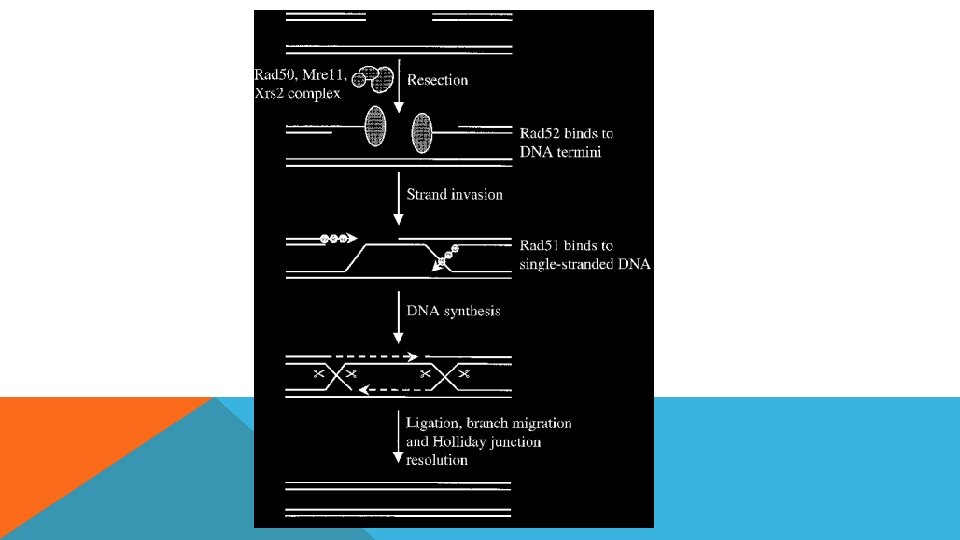
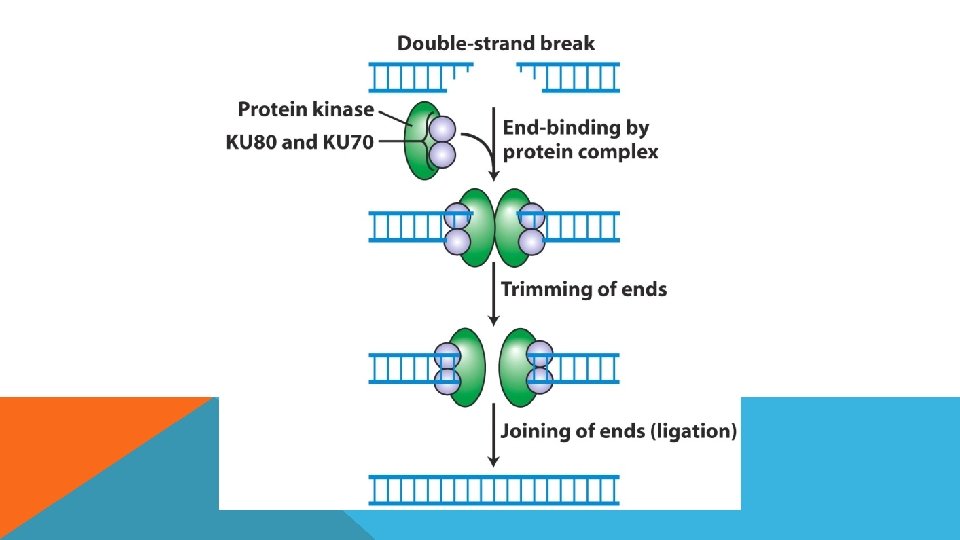
 POLİMERİZASYON")
ERROR-PRONE (HATA EĞİLİMLİ) POLİMERİZASYON

HASTALIKLAR • Kolon Kanseri • Hücresel UV duyarlılığı • Werner Sendromu (erken yaşlanma, büyümede gecikme) • Bloom Sendromu (günışığına duyarlılık) • Xeroderma pigmentosum (günışığına aşırıduyarlılık, erken yaşta deri kanseri) • Cockayne sendromu
- Slides: 95