DNA Replication I u Any eukaryotic chromosome contain
 王之仰")
")
 the two ends, or")
,")
 leads to a equal or nearly equal")
 sequences ligated to their ends. u Three regions (I, II, and")

















 became significantly shorter; the defects included depletion of tissues that require high rates")


- Slides: 38
DNA Replication (I) 王之仰
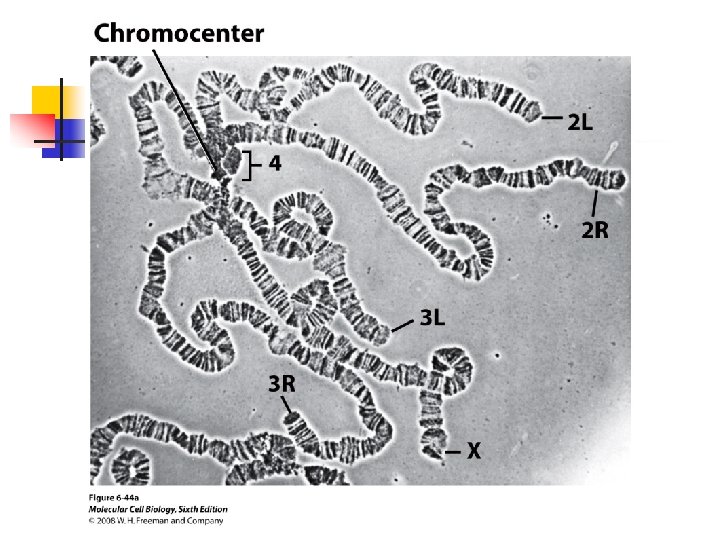
u Any eukaryotic chromosome contain three functional elements to replicate and segregate correctly: (1) replication origins at which DNA polymerases and other proteins initiate synthesis of DNA; (2) the centromere, the constricted
region required for proper segregation of daughter chromosomes; and (3) the two ends, or telomerases. u Replication of DNA begins from sites that are scattered throughout enkaryotic chromosomes.
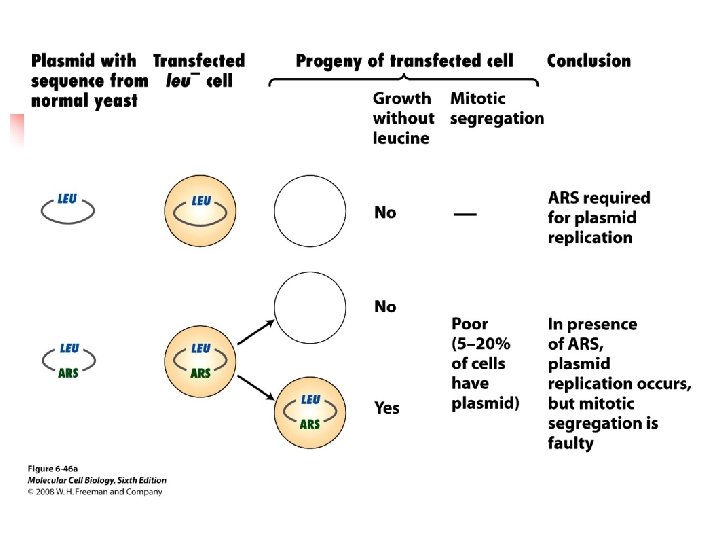
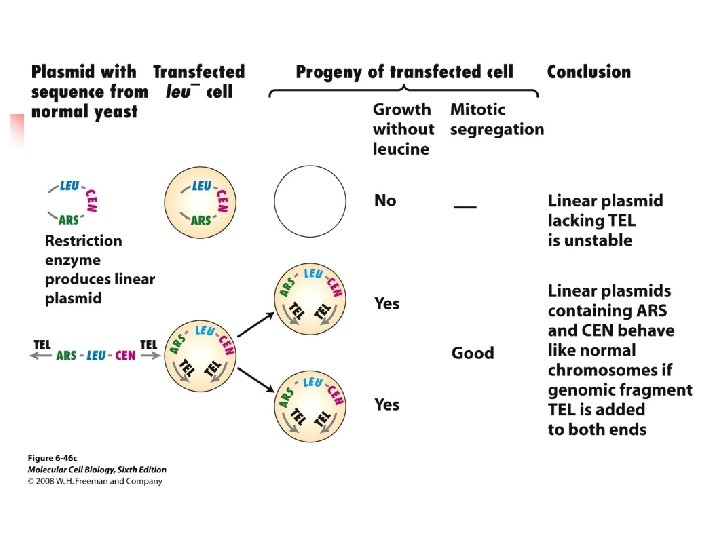
u The yeast genome contains many ~100 bp sequences, called autonomously replicating sequences (ARSs), that act as replication origins. u Up to 20% of progeny cells are faulty on the mitotic segregation.
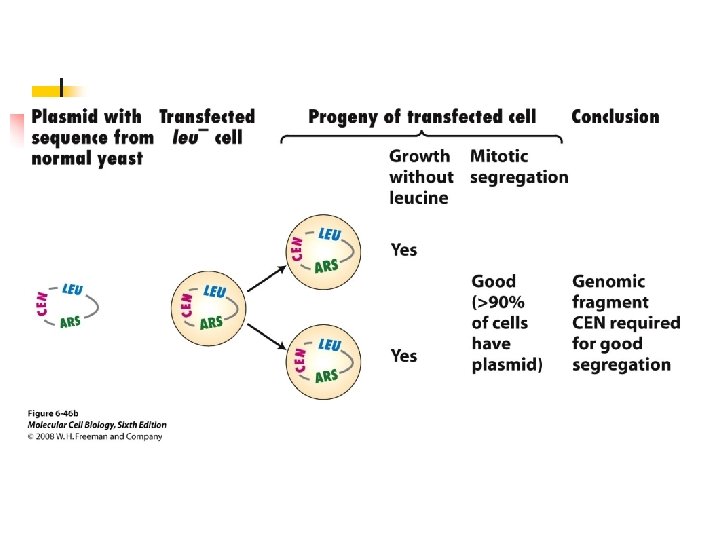
u A CEN sequence (yeast centromere sequence) leads to a equal or nearly equal segregation of yeasts during mitosis. u If circular plasmids containing an ARS and CEN sequence are cut, the resulting linear plasmids do not replicate unless they contain
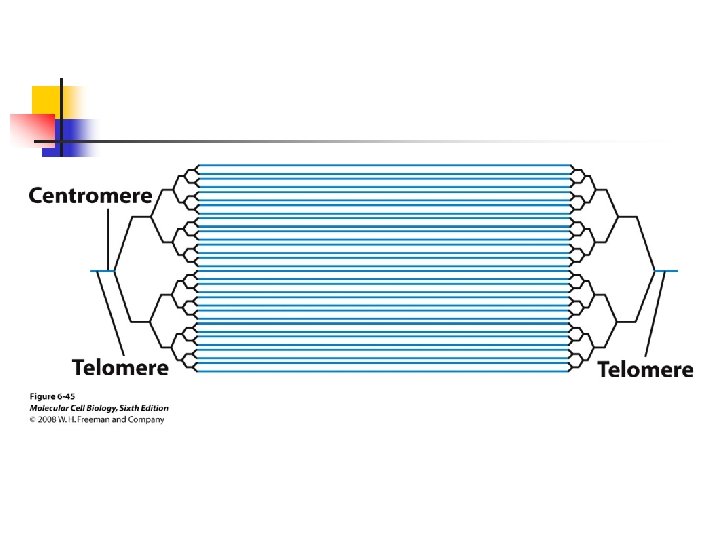
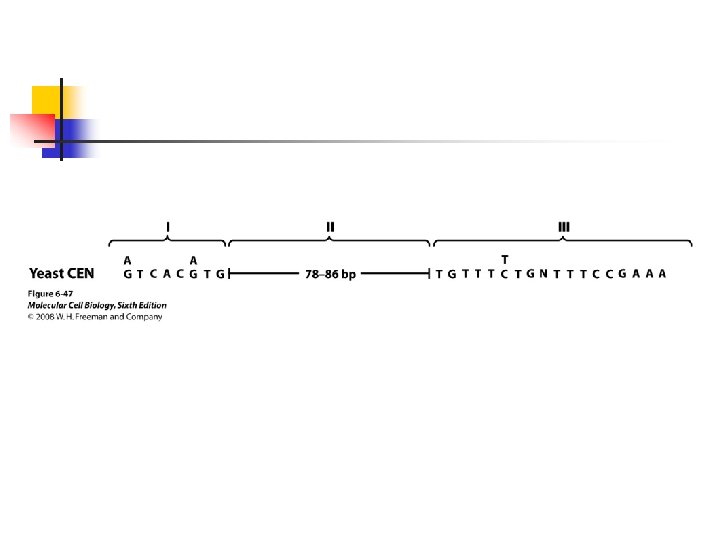
special telomeric (TEL) sequences ligated to their ends. u Three regions (I, II, and III) of the centromere are conserved among different chromosomes. u Conserved sequences are present in the region I and III.
; the region II has a fairly constant length (rich in A and T residues) and it contains no definite consensus sequences. u Regions I and III are bound by proteins that interact with more than 30 proteins.
which in turn bind to microtubules. u Region II is bound to a nucleosome that has a variant form of histone H 3 replacing the usual H 3. u Centromeres from all eukaryotes similarly are bound by nucleosomes
with this specialized, centromerespecific form of histone H 3, called CENP-A. u In human, centromeres contain 2 to 4 -megabase arrays of a 171 -bp simple-sequence DNA called alphoid DNA.
u The telomere repeat sequence in vertebrates is TTAGGG; these simple sequences are repeated at the very termini of chromosomes. u The 3’end of the G-rich strand extends 12 -16 nucleotides beyond the 5’-end of the complementary Crich strand; The region is bound
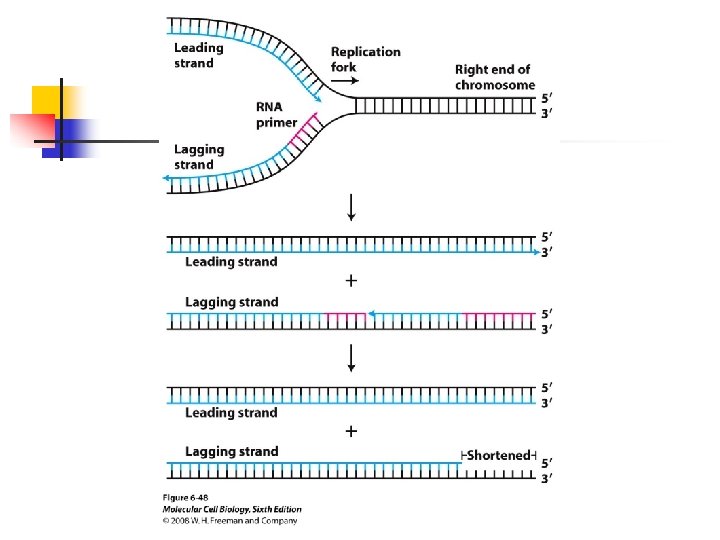
by specific proteins that protect the ends of linear chromosomes from attacked by exonucleases. u The need for a specialized region at the ends of eukaryotic chromosome is apparent when we consider all DNA polymerases elongate DNA chain at the 3’-end, all require an
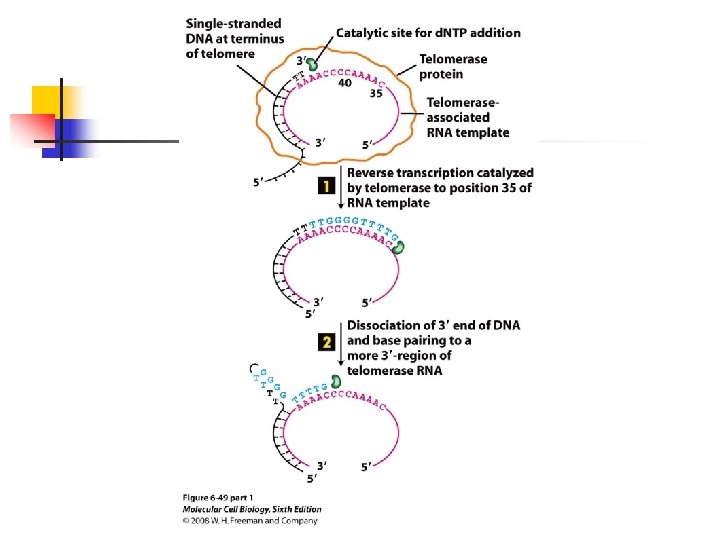
RNA or DNA primer. u Unlike the leading strand, the lagging-strand template is copied in a discontinuous fashion, it cannot be replicated in its entirety. u The telomere shortening is solved by an enzyme that adds telomeric (TEL) sequences to the ends of each
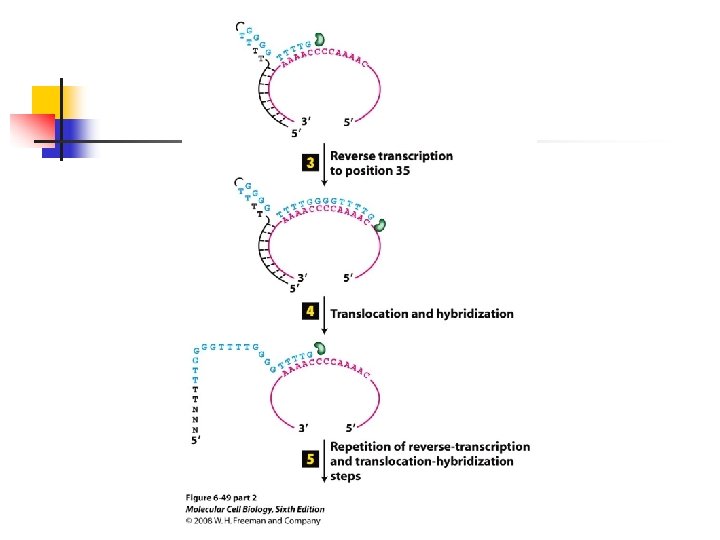
chromosome. u Because the sequence of the telomerase-associated RNA serves as the template for addition of d. NTPs to the ends of telomeres-the
source of the enzyme and not the source of the telomeric DNA primer determines the sequence added. u Telomerase is a specialized form of a reverse transcriptase that carries its own internal RNA template to direct DNA synthesis.
u The human genes expressing the telomerase protein and the telomerase-associated RNA are active in germ and stem cells, but are nearly turned off in most cells of adult cells. u These genes are activated in most cancer cells, where telomerase is
required for the multiple cell divisions necessary to form a tumor. u Telomerase prevents telomere shortening in most eukaryotes, some organisms use alternative strategies; Drosophila species maintain telomere lengths by the regulation insertion of non-LTR
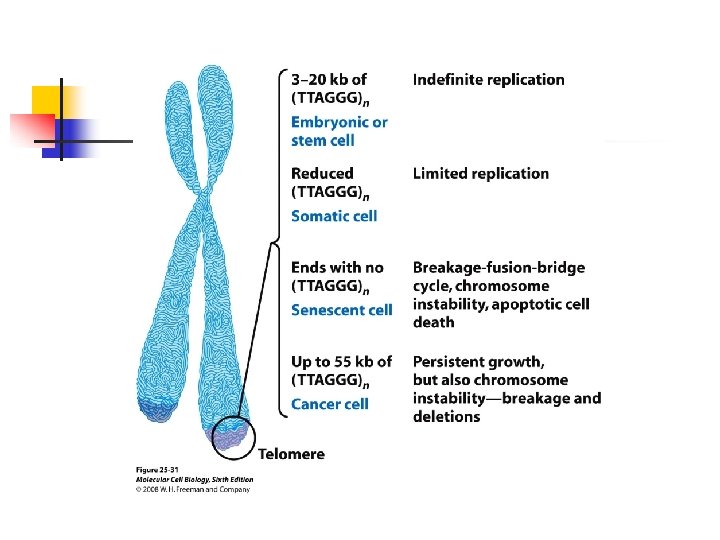
retrotransposons into telomeres. u Telomeres: the physical ends of linear chromosomes, consist of tandem arrays of a short DNA sequence, TTAGGG in vertebrates. Telomeres provide the solution to the end-replication problemthe inability of DNA polymerases to
completely replicate the end of a double-stranded DNA molecule. u Embryonic cells, germ-line cells, and stem cells produce telomerase, but most human somatic cells produce only a low level of telomerase as they enter S phase;
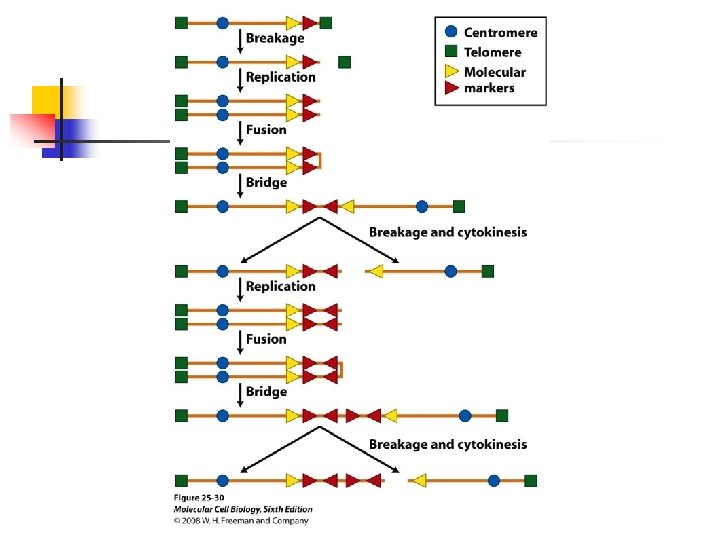
their telomeres shorten with each cell cycle. u Complete loss of telomeres leads to end-to-end chromosome fusions and cell death. u Extensive shortening of telomeres is recognized by the cell as a kind of DNA damage, with consequent
stabilization and activation of p 53 protein, leading to p 53 -triggered apoptosis. u Most tumor cells, despite their rapid proliferation rate, overcome this fate by producing telomerase. u Specific inhibitors of telomerase have been used as a cancer therapeutic
agents. u Introduction of telomeraseproducing transgenes into cultured human cells can extend their lifespan by more than 20 doublings while maintaining telomere length. u Treating human tumor cells with anti-sense RNA against telomerase
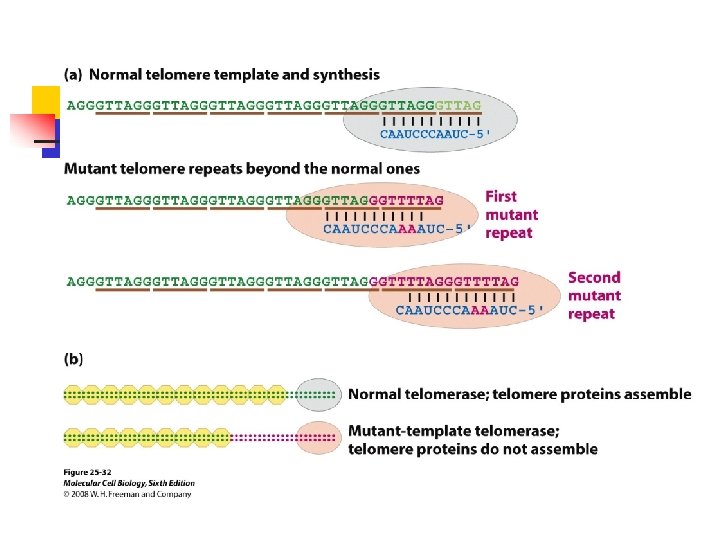
caused them to cease growth in about four weeks. u Dominant-negative telomerases, such as those carrying a modified RNA template, can interfere with cancer cell growth-when such a mutant was expressed in prostate or breast cancer cells, the cells became apoptotic.
u Genetic approaches have demonstrated that mice homozygous for a deletion of the RNA subunit of telomerase are viable and fertile was surprising. However, after four to six generations defects began to appear in the telomerase-null mice as their very long telomeres (40 -60
kb) became significantly shorter; the defects included depletion of tissues that require high rates of cell division, like skin and intestine, and infertility. u Skin papilloma induced by a combination of chemical carcinogens occurs 20 times less
frequently in mice lacking a functional telomerase than in normal mice, presumably because p 53 -triggered apoptosis is induced in response to the ever shortening telomeres of cells that have begun to divide. u If both telomerase and p 53 are absent, there is an increased rate of epithelial
tumors such as squamous-cell carcinoma, colon, and breast cancer. u Mice with an APC mutation normally develop colon tumors, and these too are reduced if the mice lack telomerase.