DISANJE Istorijat prouavanja procesa disanja prouavanja procesa disanja

 riješio problem odnosa života")




































")





 predstavlja odnos između izdvojenog CO 2 i usvojenog O")






- Slides: 53
DISANJE Istorijat proučavanja procesa disanja - proučavanja procesa disanja potiču još iz praistorijskog doba, a prvi zapisi se javljaju prije više od hiljadu godina----zapisi u Bibliji, zapisi starih Grka - terminološki pojam » disanje « se poistovjećivao sa pojmom » psiha «, » duša « ili » život « označavajući tako da je disanje znak života (onog momenta kada čovijek prestane da diše on je mrtav) – u tom periodu smatralo se da je disanje svojstveno jedino čovijeku i životinjama - egzaktija proučavanja procesa disanja javljaju se u XVII vijeku: - Robert Hook (1665) – teorija da vazduh potpomaže sagorjevanje u životinjskom organizmu - Priestley (1774) - biljke popravljaju vazduh (eksperiment je pokazao da se u zatvorenom prostoru pod staklenim zvonom svijeća gasi a da se životinje guše od » fiksiranog « vazduha; vazduh se može » popraviti « ako se pod zvono unese zelena biljka)- kiseonik nije bio poznat, Pristley ga je nazvao » deflorgistički vazduh «
-sa istorijskog stanovišta, može se reći da je Ingen-Housz (1779) riješio problem odnosa života biljke i vazduha -pokazao je da je disanje pojava koja se javlja u svim djelovima biljke, izuzev zelenih u uslovima jake osvjetljenosti-zaključio je da se disanje javlja u slučaju kada se odstrani » deflorgistički « vazduh i da se oslobađanjem » fiksiranog « vazduha (CO 2) dolazi do zamućenja i pojave bijelog taloga u karbonatnoj vodi -nije znao da li ovaj proces odvija istovremeno sa » prečišćavanjem « vazduha -Lavoisier (1789) je utvrdio da se u procesu sagorjevanja koristi vazduh koga je nazvao » kiseonik «, čime je zamjenjen dotadašnji naziv » deflorgistički vazduh «, a da » fiksirani vazduh « sadrži ugljenik i kiseonik, koji je nazvan ugljena kiselina -zaključio je da disanje životinja predstavlja sporo sagorjevanje ugljenikovih supstanci u tijelu koje je u osnovi slično sagorjevanju ugljenika---pri ovom sagorjevanju koristi se kiseonik, a oslobađaju ugljena kiselina i toplota (oslobođena toplota u procesu disanja nadoknađuje izgubljenu toplotu iz tijela)
- u XIX i početkom XX vijeka uglavnom je istraživana hemija i biohemija disanja - u procesu disanja nagomilava se energija, život biljaka zavisi od ovog procesa -upoznat je proces vrenja tj. fermentacije – utvrđeno je da je ovaj proces veoma važan u prirodi i da je vezan za životnu aktivnost ćelija; - Pfeffer je koristio izraz fermentacija u smislu anaerobnog procesa i smatrao ga je procesom disanja jer se u njemu oslobađa energija; - utvrđeno je da se fermentacija (proizvodnja alkohola) javlja u uslovima kada se vrijednost koncentracije kiseonika praktično približi nuli - dokazano je da u procesima fermentacije i disanja učestvuju različiti enzimi - kao supstrat za proces disanja mogu da se koriste različiti supstrati i njihovi međuprodukti ne moraju biti razloženi do krajnjih produkata CO 2 i H 2 O, već mogu poslužiti kao prekursori za sintezu drugih jedinjenja-----definisan je proces glikolize i trikarbonskih kiselina - utvrđeno je da ATP ima važnu ulogu u pojedinim reakcijama alkoholnog vrenja i procesima oksidacije ugljenih hidrata ili masti
- organska jedinjenja izgrađena u fotosintezi koriste se u biljci: - za rastenje, - za izgrađivanje novih ćelija i organa - za obrazovanje rezervi -disanje predstavlja proces oksidacije organskih jedinjenja u kome se oslobađa energija potrebna za obavljanje različitih funkcija biljke -postoji velika sličnost među biljkama i životinjama u pogledu biohemijskih procesa od kojih se sastoji disanje -ona ukazuje na filogenetsku starost ovih procesa - biljke se od životinja razlikuju po tome što kod njih ne postoje pokreti disanja, niti posebni organi pomoću kojih se obavlja razmjena gasova sa spoljašnjom sredinom (O 2 od svih biljnih ćelija dolazi difuzijom, a CO 2 se na isti način izlučuje, tako da se pod disanjem biljaka podrazumjeva samo ćelijsko disanje)
- kao supstrat disanja najčešće se koriste: - ugljeni hidrati i - masti - rezervni proteini mogu neki put da se koriste u disanju, ali je po pravilu ova pojava znak gladovanja -bez obzira na supstrat razlikuju se sledeće faze disanja: - pripremna faza ili faza mobilizacije, u kojoj dolazi do razlaganja složenih jedinjenja i do povišenja energetskog nivoa supstrata; - faza oksidativne degradacije supstrata, u kojoj se raskidaju veze među ugljenicima organskih jedinjenja, u kojoj se mnogi ugljenikovi atomi oksiduju do CO 2, a različiti koenzimi redukuju - faza ponovne ili terminalne oksidacije ovih koenzima, koja se sastoji u prenosu elektrona na kiseonik, pri čemu nastaje voda - oksidativna fosforilacija tj. sinteza ATP, koja je skopčana sa oksidacijom koenzima - disanje je oksidacija glukoze do CO 2 i redukcija O 2 do H 2 O C 6 H 12 O 6 + 6 O 2 ----- 6 CO 2 + 6 H 2 O
UGLJENI HIDRATI KAO SUPSTRAT DISANJA Sinteza rezervih ugljenih hidrata -vjerovatno najrasprostranjeniji oblik rezervih ugljenih hidrata je skrob (polimer -Dglukopiranoze; amiloza + amilopektin -sinteza primarnog skroba u hloroplastima - u mnogim drugim organima postaje od saharoze koja se do njih transportuje floemom, a zatim ulazi u sve ćelije tkiva za magacioniranje (u ovim ćelijama proplastidi se razvijaju u amiloplaste, u kojima se obavlja sinteza skroba) - u citosolu ćelija za magaciniranje saharoza se razlaže na dva moguća načina:
I - pomoću invertaze se razlaže na glukozu i fruktozu - zatim dolazi do fosforilacije glukoze i fruktoze da bi ušle u amiloplast II – pomoću saharozne sintaze i uridin difosfata (UDP) pri čemu nastaje uridin difosfoglukoza (UDPG) i fruktoza - UDPG pomoću pirofosfata (PPi) i enzima pirofosforilaze prelazi u glukozu-1 fosfat (G-1 -P) i UTP - G-1 -P ulazi pomoću posebnog translokatora u amiloplast i pomoću ADPglukozne pirofosforilaze prelazi u ADPglukozu koja služi za sintezu skroba - korišćenje PPi za fosforilaciju čini prvu značajnu razliku između biljnih i animalnih ćelija
Mobilizacija ugljenih hidrata za disanje u pripremnoj fazi disanja polisaharidi se najprije razlažu na monoze, koje se zatim fosforilišu da bi mogle biti oksidovane - najvažniji supstrati kod biljaka su skrob i saharoza - krajnji produkt pripreme za disanje je fruktoza 1, 6 -bisfosfat (monosaharidi koji se dobiju razlaganjem drugih ugljenih hidrata se u pripremnoj fazi prvo transformišu u glukozu ili fruktozu, koje se zatim fosforilišu) - postoje dva alternativna puta u oksidativnoj degradaciji šećera - glikoliza – oksiduje se dva puta fosforilovana fruktoza oksidativni pentozni ciklus –tzv. direktna oksidacija – oksiduje se jedanput fosforilovana glukoza
GLIKOLIZA – proces razgradnje i oksidacije šećera od F 1, 6 -BF do piruvata - pripremna faza (od skroba i saharoze do F 1, 6 -BF) -od fruktoze 1, 6 –bisfosfata do piruvata – - F 1, 6 -BP pod dejstvom aldolaze se razlaže na dvije trioze 3 -GAP i DHAP -ove trioze su u ravnoteži preko enzima triozafosfatne izomeraze (u daljim reakcijama troši se samo 3 -GAP, a njegov gubitak se nadoknađuje transformacijom DHAP, tako da na kraju obje polovine F 1, 6 -BP bivaju iskorišćene - 3 -GAP se oksiduje – to je jedina oksidativna reakcija u glikolizi (enzim gliceraldehid-3 -fosfatna dehidrogenaza, čiji koenzim NAD prelazi pritom u NADH); oksidacija aldehida u kiselinu je praćena oslobađanjem energije koja aktivira neorganski fosfat, tako da se on vezuje makroenergetskom vezom za supstrat i dobija se 1, 3 -bisfosfoglicerinska kiselina
- 1, 3 –bisfosfoglicerinska kiseline se oslobađa jedne fosfatne grupe, nju prima ADP i prelazi u ATP (enzim fosfogliceratna kinaza) i nastaje 3 - fosfoglicerinska kiselina (3 -PGA) -3 -PGA pod dejstvom fosfogliceratne mutaze daje 2 -fosfoglicerinsku kiselinu - na 2 -PGA djeluje enolaza oduzimajući molekul vode i nastaje fosfoenolpirogrožđana kiselina (PEP), enzim piruvat kinaza oduzima fosfat i dobija se pirogrožđana kiselina i ATP--kraj glikolize - neke od reakcija se sreću u reduktivnom pentoznom ciklusu (u istom ili obrnutom smjeru)
- balans glikolize glukoza + 2 ADP + 2 Pi + 2 NAD ----- 2 piruvata + 2 ATP + 2 NADH (na jedan molekul glukoze proizvode se 4 molekula ATP, ali se 2 troše za fosforilaciju fruktoze, pa je zato čist prinos cijelog procesa 2 ATP)
- energija procesa oksidacije 3 -GAP je sačuvana u redukcionom ekvivalentu NADH i u fosfatnim vezama ATP-----pošto je u ovom procesu nastajanje ATP povezano direktno sa oksidacijom supstrata proces se zove supstratna fosforilacija -u biohemiji se smatra da je PPi uzgredni produkt anabolizma i da je njegova dalja sudbina samo razlaganje na dva neorganska fosfata - ali, biljke imaju jedinstvenu sposobnost da koriste i ovu visokoenergetsku fosfatnu vezu, čime štede ATP –tzv. zaobilazne » bypass « reakcije ( kada nema dovoljno fosfata u zemljištu i kada se adenilni nukleotidi štede za druge procese u kojima su nezamjenjivi)
- veza glikolize sa drugim procesima – -glikoliza je univerzalan proces kod svih živih organizama, što ukazuje na njenu filogenetsku starost---sve reakcije su identične u svim ćelijama - međutim, različiti organizmi (bakterije, kvasci, biljke , životinje) se razlikuju po načinu na koji se koriste produkti glikolize, po načinu regulacije i po mjestu u ćeliji na kome se dešavaju - i kod biljaka to je osnovni katabolički proces koji se odvija u anaerobnim uslovima, ali se mnogo više koristi za dobijanje intermedijernih jedinjena koja služe kao početni elementi za druge sintetičke procese: - glukoza i fruktoza - njihovi fosforilovani derivati - 3 -PGA - PEP - piruvat - cio proces glikolize jer se često odvija u suprotnom smjeru i dovodi do izgradnje šećera -prije svega glikoliza se u biljnim ćelijama dešava u citosolu, kada je supstrat saharoza, a i u plastidima (u amiloplastima i u hloroplastima u mraku) -iako su ovi procesi biohemijski identični, njih obavljaju posebne grupe izoenzima, koje čak kodiraju i posebni geni---tako se u istoj ćeliji mogu dešavati 2 glikolitička procesa paralelno, u istom ili suprotnom smjeru, , , međuprodukti se razmjenjuju translokatorima na membrani plastida, ali procesi ne moraju biti regulisani na isti način
Fermentacija - pirogrožđana kiselina, dobijena procesom glikolize može biti korištena na dva načina, što zavisi od spoljašnjih uslova: - u anaerobnim uslovima, dolazi do fermentacije ili vrenja, - u aerobnim uslovima, u prisustvu O 2 pirogrožđana kiselina se oksiduje u procesu “ciklus trikarbonskih kiselina” - NADH koji je nastao u glikolizi ne može da se oksiduje, pa bi njegovo nagomilavanje u reduktivnoj formi moglo da dovede do zaustavljanja procesa-----NADH se oksiduje ponovo na dva načina -alkoholno i -mliječno vrenje - alkoholno i mliječno vrenje su karakteristični procesi za neke mikroorganizme (upotreba kvasnih gljivica koje vrše alkoholno vrenje u industriji hleba i piva; upotreba bakterija Lactobacillus u preradi mlijeka) - više biljke ne mogu dugo da žive pod anaerobnim uslovima (sjemena u prvim fazama klijanja jer O 2 ne prolazi kroz sjemenjaču ili nemaju enzime za aerobno disanje; krtole u središnjim djelovima, korjen; pirinač u ranoj fazi razvića)
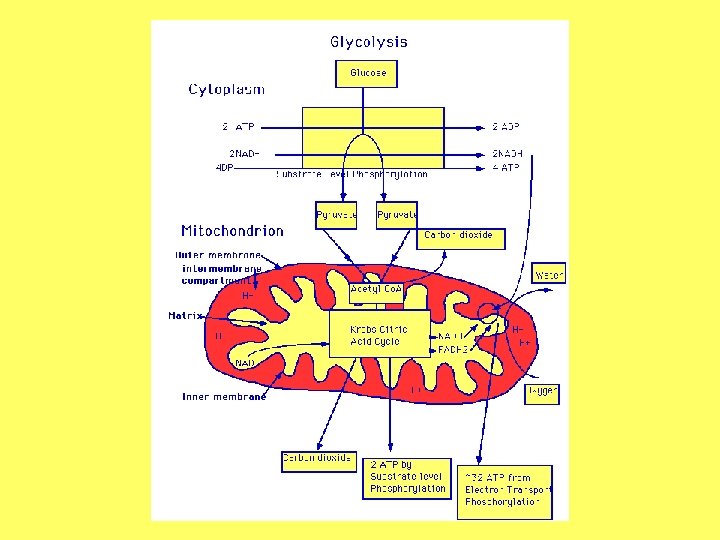
CIKLUS TRIKARBONSKIH KISELINA –ciklus linunske kiseline – Krebsov ciklus -drugi stupanj disanja, TCA ciklus obavlja se kod svih živih bića u matriksu mitohondrija (kod biljaka i u mikrotjelima) Od piruvata do acetil-Co. A - piruvat koji nastaje u citoplazmi kao krajnji produkt glikolize, prolazi kroz unutrašnju membranu mitohondruja pomoću proteinskog translokatora koji razmjenjuje piruvat za OH- jon - kompleks od tri enzima pod imenom piruvat dehidrogenaza učestvuje u prevođenju piruvata u acetil-Co. A koji ulazi u Krebsov ciklus
Oksidacija organskih kiselina - acetil-Co. A sadrži tioestarsku vezu bogatu energijom, za račun koje se kondezuje sa oksalacetatom pomoću limunske sintaze, a Co. A se ponovo odvaja----- proizvod je limunska kiselina ----prva trikarboksilna po kojoj je ciklus dobio jedan od naziva - citrat gubi vodu (enzim akonitaza) i prelazi u izocitrat—promjena OH grupe sa 3 na 4 C atom - izocotrat se oksiduje u oksaloćilibarnu kiselinu (izolimunska dehidrogenaza, koenzim NAD) koja ostaje vezana za enzim i gubi CO 2 i postaje -ketoglutarna kiselina
- oduzima se CO 2, pomoću lipoične kiseline vezuje se Co-A i dobija sukcinil-Co-A - pomoću sukcinatne tiokinaze raskida se estarska veza i dobija sukcinat(ćilibarna kiselina) i Co. A, a oslobođena energija tiestarske veze se koristi za sintezu ATP (kod animalnih mitohondrija tu se obrazuje GPP) - sukcinat se oksiduje u fumarat (koenzim flavin adenin dinukleotid FAD-ne nalazi se u matriksu mitohondrija nego je vezan za unutrašnju membranu i pripada u isto vrijeme elektron transportnom lancu) - fumarna kiselina prima vodu (enzim fumaraza) i nastaje malat - malat se oksiduje (enzim jabučna dehidrogenaza, koenzim NAD) u oksaloacetat
- bilans TCA ciklusa: - 3 C ulaze u ciklus kao piruvat, a pojavljuje se 3 CO 2 2 PIRUVATA + 8 NAD + 2 FAD + 2 ADP + 2 Pi---6 CO 2 + 8 NADH + 2 FADH + 2 ATP
Veza TCA ciklusa sa drugim procesima - prije svega TCA je povezan sa glikolizom i oba čine ne samo dio glavnog oksidativnog procesa u ćeliji, nego služe za proizvodnju intermedijernih jedinjenja koja su početni supstrat u drugim biosintetičkim putevima
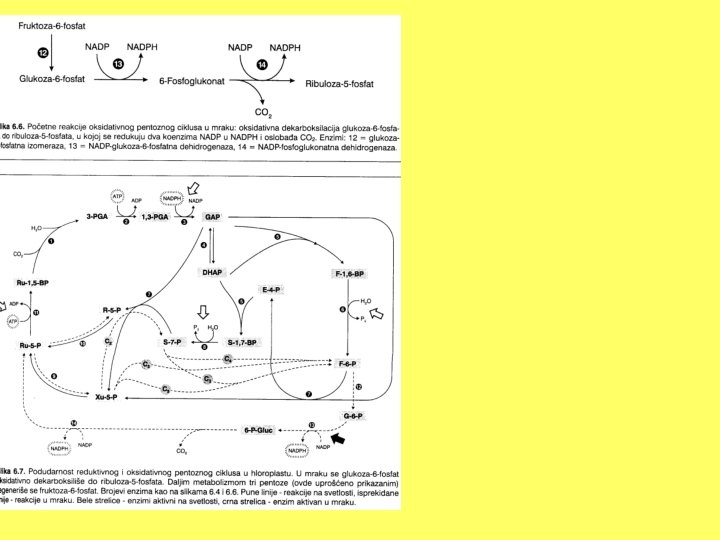
OKSIDATIVNI PENTOZNI CIKLUS - direktna oksidacija glukoze je alternativan put za oksidaciju ugljenih hidrata - sastoji se iz dvije faze: - oksidativne (koja je ireverzibilna) i - reverzibilne faze koja obuhvata metabolizam pentoza - ovaj proces se obavlja u citosolu i u hloroplastima u mraku (na svjetlu reduktivni pentozni ciklus, a u mraku niz reverzibilnih reakcija koje predstavljaju oksidativno pentozni ciklus)
Oksidativna faza - G-6 -P se oksiduje u 6 -P-glukonat (glukoza-6 -fosfatna dehidrogenaza, koenzim NADP) - oksidativnom dekarboksilacijom 6 -P-glukonata (6 -fosfoglukonatna dehidrogenaza, koenzim NADP) dobija se ribuloza-5 -P - ovo su jedine dvije oksidativne reakcije u ciklusu i obje daju po jedan redukovani koenzim NADPH
Rekombinacija pentoza - ribulozo-5 -P ulazi u niz reakcija u kojima se regeneriše početno jedinjenje G-6 -P (reakcije su suprotnog smjera od onog u reduktivno pentoznom ciklusu) -Ru-5 -P prelazi u ksilulozu-5 -P (ribulozafosfatna epimeraza) i u ribozu-5 -P (riboza fosfatna izomeraza)---ove tri pentoze su u ravnoteži - transketolaza čiji je koenzim TPP (tiamin piro fosfat) odvaja od Xu-5 -P dvokarbonski fragment i kondezuje sa R-5 -P i nastaje S-7 -P-------a kao ostatak 3 -GAP - transketolaza odvaja od S-7 -P fragment od 3 ugljenika, spaja sa 3 -GAP i daje F-6 -P - E-4 -P se spaja sa još jednim dvokarbonskim fragmentom vezanim za TPP i ponovo gradi F-6 -P - izomerazom F-6 -P prelazi u G-6 -P
Veza oksidativnog pentoznog ciklusa sa drugim procesima -samo mali broj ugljenikovih atoma se oksiduje u CO 2, a ostatak služi za regeneraciju glukoze -energija oksidativnih reakcija je sačuvana u obliku redukovanih koenzima koji se ređe ponovo oksiduju u terminalnoj oksidaciji, već obično služe u ćeliji kao reduktanti u mnogim drugim reakcijama -pentozni ciklus ima vrlo veliku ulogu u obrazovanju reduktanata u citoplazmi, što je od značaja za nefotosintetičke ćelije, za tkiva u diferenciranju, za klijanje sjemena, kao i u toku mračnog perioda u hloroplastu -proizvodi fosforibozu koja je potrebna za sintezu nukleotida i NK -jedan od intermedijera, koji se rijetko javlja u drugim procesima, eritroza-4 -P---poslije kondezacije sa PEP daje predursore za sintezu aromatičnih kiselina (triptofana, tirozina i fenilalanina---kod biljaka triptofan je prekursor auksina, a od tirozina i fenilalanina nastaju mnoga sekundarna jedinjenja kao što su fenolna jedinjenja, pigmenti, lignin. . . )---više povezan sa sintetičkim nego sa degradativnim procesima u ćeliji
- u tijesnoj vezi sa glikolizom: - oba procesa se javljaju uporedo u dva ćelijska kompartmenta –u hloroplastu i u citosolu - povezuju ih trioze 3 -GAP i DHAP -u glikolizi su postanak i konverzija trioza reverzibilni sve do PEP, a u pentoznom procesu su svi procesi, izuzev početnih oksidacija, takođe reverzibilni -slobodna razmjena trioza između ova dva procesa omogućava balans između drugih, sekundarnih biosintetičkih puteva - tako korišćenje riboza-5 -fosfata za sintezu NK iscrpljuje pul pentoza u pentoznom ciklusu ali se nedostatak popunjava dotokom trioza iz glikolitičkog puta, koje se na odgovarajućem mjestu uključuju -takođe, pojačana sinteza aromatičnih AK, u kojoj se troše eritroza-4 -fosfat i PEP, mogla bi da zaustavi neki od procesa, kada se trioze nebi mogle prelivati iz jednog u drugi -ireverzibilne oksidativne reakcije od glukoza-6 -fosfata se koriste ubrzano kada se NADPH troši u drugim reduktivnim procesima
LIPIDI KAO SUPSTRAT DISANJA -lipidi – grupa jedinjenja u koju ulaze supstance različite strukture – svrstave su u istu kategoriju uglavnom na osnovu svoje nerastvorljivosti u vodi - ekstrahuju se iz biljnog materijala organskim rastvaračima (metanol, aceton, hloroform, i dr. ) - po ovoj def. , među lipide spadaju mnoga izoprenoidna jedinjenja, kao steroli i karotenoidi, zatim jedinjenja čiji je samo jedan dio izoprenoidnog porijekla, kao hlorofil i kinoni - zajednička osobina većine lipida je osnovni gradivni dio njihovog molekula porijeklom od acetil-Co. A
-strukturni lipidi – ulaze u sastav bioloških membrana i voskovi – grade zaštitni sloj na površini mnogih organa -rezervni lipidi – nakupljaju se u sjemenima i plodivima ( u sferozomima) - biljni lipidi su veoma važan sastojak u ishrani sisara – u ćelijama sisara se ne izgrađuju linolna i linoleinska nezasićena masna kiselina (bez njih ne može da se izgrade ćelijske membrane) – ove dvije kiseline su esencijalne za sisare i moraju se unijeti u organizam hranom -biosinteza lipida obuhvata nekoliko stupnjeva i raspoređena je u različitim organelama - oksidativna degradacija lipida je proces djelimično sličan sintezi, samo suprotnog smjera, a takođe se dešava u različitim organelama
Biosinteza rezervnih lipida -po sastavu svi rezervni lipidi su trigliceridi tj. estri glicerina i masnih kiselina - glicin nastaje od gliceraldehid-3 -fosfata - biljne masne kiseline su nerazgranati lanci, sa jednom karboksilnom grupom, obično imaju paran broj C (12 -20) i mogu biti zasićene, ili sadrže 1 -3 nezasićene veze
- obavlja se isključivo u plastidima (listhloroplasti; u ćelijama koje nisu zelene – proplastidi, etioplasti, leukoplasti i hromoplasti) – samo plastidi sadrže protein ACP (acyl carrier protein) –protein-acilni prenosilac Biosinteza masnih kiselina Malonil-Co. A - acetil-Co. A nastaje u mitohondrijama oksidativnom dekarboksilacijom piruvatamembrana nije propustljiva za njega, ali se oslobađa acetat koji se transportuje u plastide gdje se uz učešće ATP i Co. A gradi acelil-Co. A - vezivanjem CO 2 za acetil-Co. A dobija se malonil-Co. A - malonilni ostatak se prenosi na ACP koji ostaje vezan tioestarskom vezom za sva intermedijerna jedinjenja, sve do završetka sinteze masne kiseline - malonilni ostatak se dekarboksiliše i kondezuje sa još jednim acelil-Co. A i dobija se fragment sa 4 C atoma – 3 -ketobutril-ACP
- redukcija 3 ketobutiril-ACP pomoću NADPHdehidrogenaze, zatim dehidratacija, još jedna redukcija -----nastaje butiril-ACP –osnovni fragment u sintezi masnih kiselina - butiril-ACP se u svakom obrtu spirale lanac pruduži za dva ugljenika 16 C atoma - palmitinska kiselina 18 C atoma – stearinska kiselina - ako stearinska kiselina dobije jednu nezasićenu vezu oleinska kiselina -za vezivanje svakog acetilnog ostatka potrebna je energija fosfatne veze jednog ATP i redukcija pomoću dva NADPH, a pri tom se oslobađa jedan CO 2 - svi enzimi koji učestvuju u biosintezi masnih kiselina su dio kompleksa sintaze masnih kiselina, koja sadrži 7 različitih enzima i nalaze se u hloroplastima
Biosinteza triglicerida - esterifikacija glicerola se obavlja u membrani glatkog ER - masne kiseline, vezane za Co. A, izlaze iz plastida i ulaze u prostor membrane, između dva lipidna sloja - tu može doći do daljih promjena u njihovoj strukturi, kao što je produženje lanca (elongacija) i obrazovanje nezasićenih veza (desaturacija) - masne kiseline koje će ući u lipide vezuju se jedna do druge estarskom vezom za alkoholne grupe glicerola - kada se u prostoru između dva lipidna sloja membrane ER nakupi veća količina triglicerida, taj dio se otkida i formira se posebna organela – sferozom - lipidi izgrađeni u jednom ćelijskom kompartmentu ne prolaze slobodno kroz membrane – postoje posebni proteini tkz. proteini prenosioci lipida koji obavljaju transport između organela i ćelija
Oksidacija rezervnih lipida - rezervni lipidi se prije oksidacije razlažu na glicin i masne kiseline -hidroliza se dešava u sferozomu, enzim lipaza je vezana za unutrašnju površinu lipidne membrane -glicerin se u citoplazmi fosforiliše i kao 3 -GAP ulazi u glikolizu gdje se oksiduje - masne kiseline se razlažu u procesu koji je poznat kao -oksidacija čiji je produkt acetil-Co. A koji ulazi u TCA ciklus -mitohondrije svih eukariota sadrže sve enzime za -oksidaciju, ali noviji podaci ukazuju, da se ona kod biljaka obavlja najvećim dijelom, ili čak isključivo u glioksozomima i peroksizomima – u svakom slučaju kada se masti razlažu da bi se od njih izgradili ugljeni hidrati, acetil-Co. A ulazi u glioksilatni ciklus – u glioksizomima, zatim se u procesu glukoneogeneze obrazuju šećeri u saradnji glioksizoma, mitohondrija i citoplazmatičnih enzima
-oksidacija masnih kiselina - da bi masne kiseline mogle biti oksidovane, one se aktiviraju vezivanjem za Co. A (analogno oksidaciji šećera prije oksidacije); potrebna energija potiče od ATP, koji se razlaže na AMP i PPi i dobija se acil-masna kiselina - acil-masna kiselina se oksiduje pomoću acildehidrogenaze (čiji je koenzim FAD- za razliku od enzima animalnih mitohondrija, kod biljaka ovaj enzim prenosi vodonik na O 2 i obrazuje H 2 O 2 koji se razgrađuje na vodu i kiseonik) – postaje nezasićeni acil-Co. A masne kiseline –nezasićena veza između 2 i 3 C atoma - enzim enoilhidrataza hidrira ovu nezasićenu vezu i dobija se hidroksiacil-Co. A masne kiseline - enzim dehidrogenaza čiji je koenzim NAD oksiduje ovo jedinjenje i na C-3 atomu se formira keto grupa - pod dejstvom tiolaze i uz učešće još jednog molekula Co. A lanac masne kiseline se raskida kod -C-atoma i dobija se acetil-Co. A i acil masne kiseline, koji je za dva C kraći od prethodnog
- sa ovim supstratom isti niz reakcija se ponavlja i pri svakom ponavljanju odvaja se po jedan molekul acetil-Co. A, tako da proces traje sve kod se cio lanac masne kiseline na taj način ne degraduje - iako se reakcije ponavljaju, početno jedinjenje je svaki put kraće za 2 C pa se ne može govoriti o ciklusu nego o spirali -oksidacija
- proces -oksidacije kod biljaka je biohemijski vrlo sličan onome kod životinja, ali bitna razlika je u značaju koji taj proces ima za biljke i za životinje -kod životinja rezervne masti su glavni izvor energije, a mitohondrije su glavno mjesto transformacije te energije u ATP -kod biljaka -oksidacija u mitohondrijama, ako se uopšte obavlja, ima daleko manji značaj (vjerovatno se tim putem oksiduju samo strukturni lipidi u ciklusu njihove degradacije i ponovne sinteze) - kod biljaka oksidacija rezervnih ulja nije usmjerena na sintezu ATP, nego služi u izgradnji ugljenih hidrata, što je dominantan proces u sjemenima koja klijaju (u glioksizomima, a u drugim ćelijama u peroksizomima)
Glioksilatni ciklus - predstavlja modifikaciju TCA ciklusa, obavlja se u glioksizomima i peroksizomima - i u ovom ciklusu acetil-Co. A se kondezuje sa oksaloacetatom u limunsku kiselinu, a zatim nastaje izolimunska kiselina - ne dolazi do oksidativne dekarboksilacije – nego se izolimunska kiselina razlaže na glioksilat-jedinjenje sa 2 C atoma, i na sukcinat - sukcinat prelazi iz glioksizoma u mitohondriju, gdje se oksiduje u TCA ciklusu do malata - glioksilat se kondezuje sa još jednim molekulom acetil-Co. A i daje jabučnu kiselinu –malat (enzim malatna sintaza) - enzim malatna dehidrogenaza oksiduje malat u oksaloacetat i tako se regeneriše akceptor acetil-Co. A za novi ciklus - ni jedan C se ne oksiduje do CO 2 nego se štede i koriste za dalje procese
Glukoneogeneza - malat koji je nastao u mitohondriji ili glioksizomu izlazi u citoplazmu i oksiduje se u oksaloacetat - oksaloacetat u prisustvu ATP i pod dejstvom PEPkarboksikinaze se dekarboksiliše i fosforiliše i dobija se PEP - PEP ulazi u glikolitički proces, koji protiče u obrnutom smjeru i naziva se glukoneogeneza (sve reakcije u glikolizi su reverzibilne izuzev one u kojij PEP prelazi u piruvat-ulaskom direktno u reverznu glikolizu ovaj ireverzibilan korak je izbjegnut) - na kraju se izgrađuje se saharoza- to je način na koji se u sjemenima koja klijaju rezervna ulja pretvaraju u saharozu i koja se transportuje u mladu biljku i služi tek tada za sintezu gradivnog materijala i kao izvor energije - oksidacija šećera u mladoj biljci preko TCA ciklusa ili oksidativnog pentoznog ciklusa povezuje metabolizam rezervnih lipida sa sintezom mnogih drugih jedinjenja neophodnih za rastenje
TRANSPORT ELEKTRONA - ćelije sadrže ograničenu količinu koenzima, pa se mora uspostaviti ravnoteža između redukovanih i oksidovanih oblika, da bi ćelijske funkcije mogle kontinuirano da se obavljaju - koenzimi redukovani u disanju se ponovo oksiduju, prenoseći elektrone na O 2 pri čemu se obrazuje voda tzv. terminalna oksidacija - prenos elektrona se obavlja spontano niz gradijent redoks potencijala, a povezan je sa vektorskim transportom protona sa jedne na drugu stranu membrane, javlja se izvjesna količina slobodne energije koja je akumulirana kao protonska motorna sila i služi za sintezu ATP, u procesu oksidativne fosforilacije
- prenosioci elektrona su sastavni djelovi membrana mitohondrija - unutrašnja membrana se veoma razlikuje od svih drugih membrana- proteini čine 70% unutrašnje membrane, a među lipidima se nalazi kartiolipin koga nema u nijednoj drugoj membrani -unutrašnja membrana je za razliku od spoljašnje manje propustljiva - međutim mnogi proteini u njenom sastavu imaju transportnu funkciju i regulišu transport u oba pravca - pored njih glavni funkcionalni djelovi membrane su proteini koji obavljaju transport elektrona od NADH do O 2
- prenosioci elektrona su jedinjenja sa različitim redoks potencijalom: - piridin nukleotidi (NAD; NADP) - flavoproteini (FMN; FAD) - citohromi (citohrom a, b ili c) - fero-sumporna jedinjenja i - kinoni (ubikinon –koenzim Q) - svi prenosioci elektrona su integrisani proteini u unutrašnjoj membrani mitohondrija a grupisani su u 4 supramolekulska kompleksa – njih povezuju mobilni prenosioci – ubikinon u lipofilnoj središnjoj zoni membrane i - citohromi c (sa dvije komponte c 1 i c; c 1 vezan za membranu a c je asocirani protein na spoljašnjoj površini u intermembranskom prostoru
- kompleksi I-IV nisu u strukturi membrane fiksirani za pojedina mjesta - prenos elektrona opisan je kao rezultat njihove slobodne lateralne difuzije i slučajne kolizije -pravac prenosa je određen redoks potencijalom prenosilaca - najniži redoks potencijal ima par NADH/NAD koji je početni donoe elektrona u ET lancu (još jedan ulaz elektrona u ET lanac je par FADH/FAD) - oba redoks pare elektrone predaju ubikononu, koji redukuje kompleks III, zatim preko citohroma c kompleks IV i na kraju vodu
- potencijalna razlika od NADH do vode iznosi 1, 14 V -redoks potencijal para H 2 O/O 2 je 0, 81 V; -slobodna energija koja se dobije pri prenosu 2 elektrona između NADH/NAD i H 2 O/O 2 je oko 220 k. J mol-1, a između FADH/FAD i H 2 O/O 2 oko 167 k. J mol-1 -prema tome, oksidacijom sukcinata (FAD-dehidrogenaza) oslobađa se manje energije nego pri oksidaciji drugih supstrata na koje djeluju NAD-dehidrogenaze
Disanje neosjetljivo prema cijanidu - cijanid je veoma jak inhibitor za animalne mitohondrije kod kojih se svodi disanje na oko 1% od kontrolnog; cijanid izaziva trenutnu smrt životinja i čovijeka - kada se biljke izlože cijanidu potrošnja O 2 je u značajnoj mjeri spriječena ali nikada potpuno - proces u kome se troši O 2 u prisustvu CN- zove se disanje rezistentno prema cijanidu - mlada biljna tkiva su većinom potpuno inhibirana u prisustvu cijanida i ne razlikuju se od animalnih, kod većine biljaka disanje je smanjeno na 10 -25% od ukupnog disanja - postoje biljke čiji su pojedini organi potpuno neosjetljivi prema cijanidu, čak cijanid i stimuliše potrošnju kiseonika
- pokazalo se da u ET lancu motohondrija postoji enzim ubikinol oksidaza koja prenosi protone i elektrone sa ubikinola na alternativnu oksidazu ---prema tome u alternativnom putu elektroni ne odlaze na kompleks III i IV, nego skreću ranije u pravcu terminalne oksidacije -značaj disanja neosjetljivog prema cijanidu za biljku nije jasan -za Araceae vjerovatno se ovaj proces koristi za termogenezu tj. za proizvodnju toplote, na višoj temperaturi iz spadiska (zadebljali organ nalik na klip na kome stoje cvjetovi) isparavaju neka jedinjenja sa jakim mirisom koji privlače insekte i obezbjeđuju polinaciju npr. kod Arum italicum temperatura dostiže i 51 0 C pri spoljašnjoj od 15 0 C - mjerenjem disanja različitih organa pokazano je da se, na primjer, u korjenu uključuje alternativni put kada on raspolaže većom količinom ugljenih hidrata, od one koju može da iskoristi - kod listova pšenice i spanaća alternativni put se ne javlja ujutro, kada je nivo ugljenih hidrata nizak, nego tek poslije nekoliko časova fotosinteze, kada se izgradi značajna količina ugljenih hidrata
Alternativne oksidaze u citoplazmi -mnoge mebrane van mitohondrija sadrže kraće elektron-transportne lance, koji oksiduju NADH ili NADPH, a redukuju različite akceptore elektrona među kojima su: citohromi, kinoni, Fe-S proteini i dr. -elektron-transportni lanci se sastoje većinom od flavoproteina i od jednog citohroma b - transport elektrona kroz ove prenosioce nije povezan sa sintezom ATP
POKAZATELJI DISANJA Koeficijent disanja (RQ) predstavlja odnos između izdvojenog CO 2 i usvojenog O 2 - oksidacija jednog molekula glukoze može se prikazati jednačinom: C 6 H 12 O 6 + 6 O 2 = 6 CO 2 + 6 H 2 O + 2822 k. J prema tome RQ za glukozu = 6 CO 2 / 6 O 2 = 1 - oksidacija jednog molekula masti (tripalmitin) može se prikazati jednačinom: 2 C 51 H 98 O 6 + 145 O 2 = 102 CO 2 + 98 H 2 O prema tome RQ tripalmitina je 102 CO 2/145 O 2 = 0, 7 - vrijednosti RQ su različite i ne zavise samo od organske materije koja se oksiduje već i od niza drugih činilaca (T, sadržaj O 2, sadržaj vode, dužina bubrenja sjemena, debljina sjemenjače. . . ) pa je i razumljivo što se javljaju razlike u RQ kod pojedinih vrsta organa i tkiva biljaka.
Intenzitet disanja – količina usvojenog O 2 ili izdvojenog CO 2 u jedinici vremena i za određenu masu biljnog materijala (obično se izražava u mg CO 2 na 1 g suve materije za 24 sata) -ova vrijednost je nepostojana, ne zavisi samo od specifičnosti vrste biljaka, već i od osobina pojedinih organa i tkiva jedne iste biljne vrste - intenzitet disanja se mijenja zavisno od niza činilaca, međutim veličina promjena zavisi od jačine dejstva određenog činioca ili kombinacije dejstva nekoliko činilaca -temperatura- disanje se povećava sa povećanjem temperature -sadržaj kiseonika- intenzitet disanja zavisi od odnosa aerobnog i anaerobnog disanja – sadržaj kiseonika pri kome počinje anaerobno disanje je različit – kod većine tkiva smanjenje sadržaja O 2 od 20% do 5 -6% skoro da ne utiče na skretanje ka anaerobiozi---međutim pri sadržaju od 3% dolazi do povećanja koeficijenta disanja
-sadržaj ugljendioksida- pri povećanju sadržaja CO 2 disanje se usporava -svjetlost- poznato je da se disanje odvija i na svjetlosti ali o njihovom odnosu ima raznih zaključaka -sadržaj vode- vazdušno suvo sjeme ima veoma mali intenzitet disanja, a neznatno povećanje vlažnosti sjemena povećava intenzitet disanja----sa povećanjem vlažnosti optimalna temperatura za njegovo disanje se smanjuje -kod drugih biljnih organa kao pravilo je utvrđeno da smanjenje sadržaja vlage do određene granice kod mladih tkiva izaziva stimulaciju disanja, dok kod starih tkiva intenzitet disanja može i da se ne izmijeni; međutim, uticaj smanjenja vlažnosti na disanje zavisi i od bioloških osobina biljaka - promjena int. dis. u toku ontogeneze biljaka- najveći inenzitet disanja imaju najmlađi biljni organi i tkiva tj. oni koji se nalaze u stanju aktivnog rasta -mineralna ishrana-
UZAJAMI ODNOS DISANJA I FOTOSINTEZE - po količini energije i po sintezi organske materije to su dva suprotna procesa - međutim, ova dva procesa imaju i sličnih osobina - voda- u fotosintezi voda se koristi kao donor vodonika pomoću koga se vrši redukcija CO 2 dok O 2 odlazi u atmosferu - u disanju O 2 iz vazduha služi za oksidaciju C iz supstrata koji služi za disanje, dok H iz vode zajedno sa H iz supstrata služi kao promotorna sila za sintezu ATP - porfirini- učešće ovih jedinjenja predstavlja centralnu kariku u lancu procesa fotosinteze, odnosno u fotolizi vode - u velikom broju slučajeva učestvuju i u završnim etapama disanja, odnosno u oksidaciji vodonika kiseonikom - razlaganje vode se vrši uz učešće Mg-proteina, a biosintezu vode katalizuje najčešće Fe-protein
-stvaranje molekula organskog jedinjenja u procesu fotosinteze kao i proces razlaganja ovog molekula prilikom disanja može da se ostvari na nekoliko načina u ćeliji---interesantno je da su neki od puteva sinteze molekula heksoze u fotosintezi po svom karakteru veoma bliski putu razlaganja heksoze u procesu disanja--sličnost nije samo u osnovnim etapama kroz koje svaki od ovih procesa prolazi, već i u katalitičkom mehanizmu pomoću kojeg se ovi procesi ostvaruju npr. ako se uporedi Calvinov ciklus i heksozofosfatni ciklus pretvaranja heksoza razlika je u tome što transformacije u procesu disanja idu u dijametralno suprotnom pravcu u odnosu na pravac transformacija u fotosintezi iz ovog primjera se vidi da su fotosinteza i disanje dvije strane jedinstvenog procesa razmjene materije i energije kod biljka
-organske materije stvorene u procesu fotosinteze koje sadrže energiju predstavljaju materijalni izvor za sve heterotrofne organizme, a i za autotrofe u periodu kada oni ne ostvaruju fotosintezu -----prema tome, u toku dana za potrebe samih biljaka u gradivnom i energetskom materijalu troši se samo mali dio stvorenih organskih jedinjenja, dok veći ostaje neiskorišćen i akumuliran u vidu rezervi -- razgradnja stabilnih rezervnih jedinjenja i njihovo uključenje u procese razmjene materije ostvaruje se uglavnom u procesu disanja -—osnovna biološka uloga disanja je obezbjeđenje ćelije u energetskom materijalu -međutim, produkti bogati energijom koji nastaju u procesu disanja stvaraju se putem transformacije organskih materija koje su se nagradile u zelenim djelovima biljke u procesu fotosinteze ---ovo je osnovna razlika disanja i fotosinteze