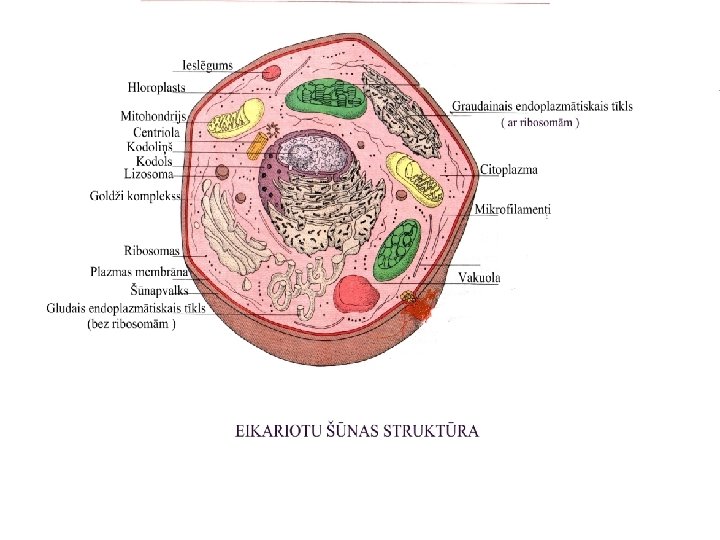
Citoplazmatisk iedzimtba Iedzimtbu ko nosaka citoplazm lokaliztie gni

Citoplazmatiskā iedzimtība Iedzimtību, ko nosaka citoplazmā lokalizētie gēni, sauc par citoplazmatisko iedzimtību.

Mitohondriju gēni Izmaiņas mitohondriju gēnos noved pie elpošanas traucējumiem - mitohondriju galvenās funkcijas.

Vislabāk mitohondriju ģenētikas pētījumiem noder raugi – saharomicētes. Raugi – saharomicētes ir fakultatīvi aerobi. Nomācot elpošanu, tie var eksistēt uz rūgšanas rēķina, izmantojot tai glikozi un dažus citus cukurus kā oglekļa avotu.

Citu, nefermantatīvu oglekļa avotu klātbūtnē, piemēram, etanola, glicerīna, kalcija laktāta u. c. , raugi neaug, ja nav elpošanas.
, pētot maizes raugu S. cerevisiae, atklāja mutantus ar")
1949. gadā Boriss Efrusi (B. Ephrussi), pētot maizes raugu S. cerevisiae, atklāja mutantus ar elpošanas nepietiekamību, kas veido sīkas kolonijas – pundurkolonijas.

Tā kā B. Efrusi ir francūzis, mazās kolonijas tika nosauktas (no franču valodas – mazs), bet normālās ( wild type )– kolonijas par (lielas).

B. Efrusi novēroja, ka petites - kolonijas bija mazas ne tāpēc, ka raugu šūnu izmēri bija mazāki, bet tāpēc, ka ievērojami samazinājās mutantā celma petites augšanas ātrums salīdzinājumā ar normālo, izejas celmu.

Petites šūnas dalās ļoti lēni, jo nesatur daudzus elpošanas cikla fermentus – citohromu a un b, citohromoksidāzi u. c. Enerģiju šīs šūnas iegūst, anaerobi fermentējot glikozi līdz etilspirtam.

• Šūnu populācijā petites- kolonijas rodas spontāni ar biežumu
 klātbūtnē, tādu kā")
Interkalējošo aģentu (vielas, kuras var iekļauties starp DNS kaimiņu bāzu pāriem) klātbūtnē, tādu kā etīdijbromīds, iespējama 100% indukcija – šūnas 100% kļūst par petite!

Tādejādi: - augsts petite rašanās biežums, - petite šūnu dzīvotspēja un augšana, iztrūkstot šūnu elpošanas funkcijām padara šo petite celmu sistēmu ērtu, lai pētītu ārpus hromosomālo iedzimtību.

Kodolu, neitrālie un supresīvie petite mutanti Dažiem petites mutantiem ir mutācija kodola genomā, kas nav brīnums, jo dažas mitohondriālo proteīnu subvienības kodē kodola gēni.
,")
Šo pundurformu nosauca par skaldīties spējīgo pundura celmu. Krustojot to ar normālo raugu (pet+), iegūtajai diploīdajai zigotai raksturīga normāla elpošana pet+/pet- un grandes kolonijas.
,")
Taču pēc sporulācijas katrā askā tikai divas no četrām sporām veidoja normālas kolonijas (grandes), pārējās divas sporas – pundurkolonijas (petites). Pašlaik ir zināmi vairāk nekā 20 kodola gēnu, kas izraisa elpošanas mutācijas.

Divas citas petites klases - neitrālie petites un supresīvie petites izrāda ārpuskodola iedzimtību. Elpošanas nepietiekamību izraisošās mitohondriālās mutācijas apzīmē [ rho-].
![– Neitrālās petites [rho-N] – [rho-N] x [rho+N] – ↓ – [rho+N] / [rho-N]](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-17.jpg "– Neitrālās petites [rho-N] – [rho-N] x [rho+N] – ↓ – [rho+N] / [rho-N]")
– Neitrālās petites [rho-N] – [rho-N] x [rho+N] – ↓ – [rho+N] / [rho-N] grandes kolonijas ↓ meiosis [rho+N] : [ rho+N] petite : grande = 0 : 4

Nosaukums neitrālās petites nozīmē to, ka šīs klases petites mutācijas neietekmē savvaļas tipu: visām paaudzēm ir viena vecāka fenotips ( Russel P. J. , 1996 )
![Neitrālo petite mutāciju daba Neitrālo [rho-] mutantu ģenētiskā materiāla izpēte parādīja pārsteidzošu faktu: 99](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-19.jpg "Neitrālo petite mutāciju daba Neitrālo [rho-] mutantu ģenētiskā materiāla izpēte parādīja pārsteidzošu faktu: 99")
Neitrālo petite mutāciju daba Neitrālo [rho-] mutantu ģenētiskā materiāla izpēte parādīja pārsteidzošu faktu: 99 - 100% trūka mt DNS. Krustojot normālos raugus, normālie mitohondriji producējas priekš visas paaudzes un petite īpašības izzūd jau pēc vienas ģenerācijas.
![Otrā petites klase, kas raksturojas ar ārpuskodola iedzimtību, ir supresīvie mutanti [rho-S]. Vairums petite](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-20.jpg "Otrā petites klase, kas raksturojas ar ārpuskodola iedzimtību, ir supresīvie mutanti [rho-S]. Vairums petite")
Otrā petites klase, kas raksturojas ar ārpuskodola iedzimtību, ir supresīvie mutanti [rho-S]. Vairums petite mutantu pieder šim tipam. Supresējošo mutantu petite pazīmju pārmantojamība atšķiras no kodolu un neitrālajiem petites.
![[rho-S] x [rho+] / [rho-S] -diploīds](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-21.jpg "[rho-S] x [rho+] / [rho-S] -diploīds")
[rho-S] x [rho+] / [rho-S] -diploīds

Mitozē pie šūnu dalīšanās diploīdajā paaudzē būs ~ 99% petites, kam nenotiek mejoze un neveidojas sporas.

Raugu diploīdam var inducēt mejozi ( to izdara vienkārši pārnesot raugus uz barotni ar kālija vai nātrija acetātu) tūlīt pēc to izveidošanās pēc supresīvo rho- šūnu un savvaļļas tipa šūnu krustošanas. Šajā gadījumā mejoze notiek normāli un ir iespējama tetrāžu analīze.
.")
Tetrāžu analīze parāda šķelšanās trūkumu lielāko daļu tetrāžu ( 0 Pet+ : 4 Pet-). (Russell P. J. , 1998 )

Mutāciju daba Izrādās, ka augsti supresīviem mutantiem saglabājas neliela mt DNS ar replikācijas sākuma punktu. Tādi mini gredzeni replicējas ātrāk nekā normālā mt DNS un ātri izspiež to pie veģetatīvās šūnu dalīšanās.

Mitohondriju ģenētika vislabāk ir izstrādāta uz raugiem – saharomicētēm, kaut gan daļa pētījumu ir veikti un rezultāti iegūti uz citiem objektiem.
 mutācijas noved pie mitohondriju morfoloģiskām izmaiņām, pie")
Mitohondriālās mutācijas Neurospora crassa Neirosporai poky (sīks) mutācijas noved pie mitohondriju morfoloģiskām izmaiņām, pie traucējumiem to olbaltumvielu sintēzē, pie atsevišķu mitohondriālo fermentu iztrūkuma. Ārēji tas izskatās pēc palēlinātas šūnu augšanas. Pazīmi manto pa mātes līniju

Neurospora crassa ir obligāts aerobs un tai ir nepieciešams skābeklis, lai augtu un izdzīvotu, tādejādi mitohondriālās funkcijas tai ir būtiskas.

Rauga sēnēm , krustojoties un veidojoties zigotām, saplūst veselas šūnas – citoplazmas un kodoli. Tas nedod iespēju, krustojot dotos mutantus ar normālu formu, atsevišķi novērtēt kā citoplazmas, tā kodola lomu pazīmju iedzimšanā.

Neirosporai iedzimtību pa mātes līniju var noteikt, pateicoties reciprokai krustošanai. Mikrokonīdijas veic vīrišķo gametu lomu, tās praktiski nepārnes citoplazmu pie apaugļošanas, sievišķo lomu veic protoperitēciji.

. Krustot A un a pārošanās tipa micēlijus var divejādi: . • . Novietot divus celmus uz barotnes vienlaicīgi. • . Barotni inokulēt ar vienu celmu un pēc 3 -4 dienām 25 C pievienot otru celmu.

Otrajā gadījumā pirmais celms producē .
, kurā viens")
Pievienojot celmu pie pretējā pārošanās tipa celma, mums ir krustojums (controlled cross), kurā viens celms kalpo kā mātes , bet otrs kā tēva vecāks.

Izmantojot celmus, kas producē protoperitēcijus kā mātes vecākus, bet cita celma konīdijas kā tēva vecākus, ģenētiķi veic reciproko (virzīto ) krustošanu. Reciprokā krustošana parāda, vai dotā pazīme ir citoplazmatiska vai hromosomāla.
![Reciprokai krustošanai ir sekojoši rezultāti: Normal♀ x [ poky] ♂→ Normal Pēc mejozes, skaldīšanās](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-35.jpg "Reciprokai krustošanai ir sekojoši rezultāti: Normal♀ x [ poky] ♂→ Normal Pēc mejozes, skaldīšanās")
Reciprokai krustošanai ir sekojoši rezultāti: Normal♀ x [ poky] ♂→ Normal Pēc mejozes, skaldīšanās būs: Normal : [ poky] = 0 : 8 [ poky] ♀ x Normal ♂ → [ poky] Pēc mejozes, skaldīšanās būs: [ poky] : Normal = 8 : 0 ( Russell P. J. , 1998 )
![Molekulārā līmenī [poky] rodas r. RNS promotora mazās mitohondriālās ribosomālās subvienības delēciju rezultātā.](http://slidetodoc.com/presentation_image_h2/0832514398c9e8cb8c3a5a059ee6d975/image-36.jpg "Molekulārā līmenī [poky] rodas r. RNS promotora mazās mitohondriālās ribosomālās subvienības delēciju rezultātā.")
Molekulārā līmenī [poky] rodas r. RNS promotora mazās mitohondriālās ribosomālās subvienības delēciju rezultātā.

Askomicētei Podospora auserina pazīstama micēlija “novecošanās“ pazīme, tas ir pakāpeniska dzīvotspējas pazemināšanas audzēšanos laikā. Dažādiem celmiem dzīves ilgums variē no 9 līdz 106 dienām. .

Ātra novecošanās tiek pārmantota pa mātes līniju

Kad normāls micēlijs saplūst ar hifu, kurai ir “novecošanas“ pazīme, normālais micēlijs inficējas.

Novecošanās pazīmes sākas tieši tajā vietā, kur saskaras normālais micēlijs ar novecojošām hifām, t. i. tieši anastomozes vietā. Attālinoties no šīm vietām, šīs pazīmes izpausmes mazinās. ( Sedzer P. , 1987 pēc Inge- Vechtomov S. G. , 1989. )

Podospora auserina hifu inficēšanās saistīta ar plazmīdām, kas infīcē mitohondrijus.
 Eksistē citoplazmatiskā iedzimtība, kas saistīta ar vīrusu un baktēriju klātbūtni")
Raugi “killer “ (killer-slepkava) Eksistē citoplazmatiskā iedzimtība, kas saistīta ar vīrusu un baktēriju klātbūtni eikariotu citoplazmā, kas atrodas simbiotiskās attiecībās ar šūnu.

Viens no piemēriem ir raugi “killer”. Daži no raugu celmiem izdala toksīnus, kas nogalina jūtīgus raugu celmus. Killer” celmi ir imūni pret saviem toksīniem.

Raugu – “killer” fenomena pamatā ir divu tipu vīrusu L un M klātbūtne šūnas citoplazmā. M vīruss atrasts tikai šūnās, kurās ir L vīruss.
")
( Rassell P. J. , 1996 )
")
L vīruss sastāv no proteīna kapsīdas un 4, 6 kb divpavedienu ( ds ) RNS genoma ( L-ds RNS ). L-ds. RNS kodē kapsīdas proteīnu gan L, gan M vīrusam, kā arī vīrusa polimerāzi, kura nepieciešama vīrusa replikācijai. Tādā veidā visas vīrusa daļiņas tiek kodētas ar L-ds RNS.

M vīruss sastāv no vīrusa daļiņas, kuru kodē L-ds RNS un genoma , kas sastāv no divām 1, 8 kb divpavedienu RNS ( M-ds RNS ).

M-ds. RNS kodē toksisko killer proteīnu, kas izdalās no šūnas. Tas pats proteīns nodrošina arī killer šūnu imunitāti.

Jūtīgās raugu šūnas tiek nogalinātas ar M- kodēto killer toksīnu. Ir divi jūtīgo šūnu tipi: viens no šūnu tipiem satur tikai L vīrusu, bet otrais – nesatur ne M, ne L vīrusus.
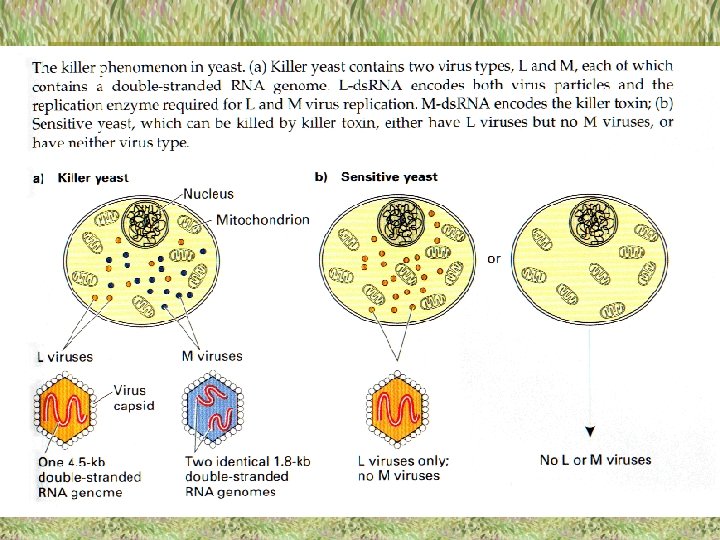

Vīrusu transmisija no šūnas šūnā notiek sajaucoties citoplazmām raugu šūnu kopulācijas laikā. Visa nākamā paaudze pārmanto vīrusu kopijas. Piemēram, killer-raugiem krustojoties ar jūtīgiem raugiem – diploīds būs killer-raugs tāpēc, ka šajā diploīdā būs L un M vīrusi no killer šūnām. Pēc mejozes no askosporām izaug raugi killer.
- Slides: 51