Chapitre 4 Gntique bactrienne Les variations gntiques par

Chapitre 4. Génétique bactérienne: Les variations génétiques par transfert de matériel génétique Jules-Roger Kuiate Professeur

Objectifs • A la fin du chapitre, l’étudiant doit pouvoir: • Expliquer les variations génotypiques et de leurs expressions phénotypiques chez les bactéries. • Justifier les conséquences des mutations en bactériologie médicale. • Identifier les différents mode de transfert génétiques, et leur implication dans la résistance au antibiotiques. • L’implication des éléments génitiques mobiles de la bactéries dans le phénomène de résistance au antibiotiques.

4. 1 Introduction • Comme tous les êtres vivants, es bactéries évoluent sans cesse dans le temps, ce qui leur permet de s’adapter à l’environnement om elles se trouvent; • Elles acquièrent ou perdent ainsi de nouvelles fonctions, par exemple métaboliques, résistance aux ATB ou augmentation de sa virulence.

• La génétique est la science qui étudie les caractères héréditaires d’un être, leur transmission de génération en générations et leurs variations (mutations). • La génétique bactérienne étudie ces caractères chez les bactéries • Génomique: étude de l’ensemble des génès d’un organisme donnée. • Les bactéries se reproduisent et conservent leurs caractéristiques de génération en génération, tout en présentant des propriétés particulières dans une faible proportion de leur progéniture.

• Gène: segment d'ADN dans lequel la séquence de bases nucléotidiques détermine üpar transcription, la séquence de bases dans une molécule d'ARNm, üpar traduction, la séquence d'AA dans un polypeptide • Mutation: altération dans la séquence de bases nucléotidiques du gène. C'est une variation : ürare : n'affecte qu'une faible fraction de l'ensemble des cellules bactériennes au sein d'une large population. üdiscontinue : ne s'effectue pas à la suite d'une longue période d'adaptation progressive, avec des formes intermédiaires, mais habituellement en une seule étape. üd'emblée héréditaire.

Notion de gène Gène: Licnce 2: Génétique - T 6 Unité fonctionnelle et physique élémentaire de l’hérédité qui transmet l’information d’une génération à la suivante. Un fragment d’ADN, constitué d’une région transcrite et de séquences régulatrices. Procaryote Eucaryote Promoteur Stop Intron Exon 1 Exon 2 Codant

4. 3 Evolution Adaptation Et Sélection Clonale • Les bactéries utilisent deux mécanismes: Mutations et le transfert génétique, mais aussi des éléments mobiles • La mutation est une modification de l'information génétique (donc ADN) dans le génome. Elle est l’une des causes principales de l‘ évolution de l’ espèce. • La mutation est : • Spontanée. • Discontinue ou brusque. • Stable.

• Rare: Il est caractéristique d'un caractère donné, de l'ordre de 10 -5 à 10 -10, le taux moyen étant de l'ordre de 10 -6. • Spécificité : la probabilité pour une bactérie de subir simultanément deux mutations distinctes est le produit des probabilités individuelles de ces mutations. Cette notion est d'importance, afin d'éviter la sélection d'un mutant résistant, dans l'antibiothérapie, antituberculeuse par exemple.

4. 3. Variations génétiques par mutation • Différents types • A. Mutations ponctuelles • Microdélétion : perte d'une paire de bases. • Microinsertion (Microaddition): gain d'une paire de bases. • Substitution: rempacement d'une paire de bases par une autre à la suite d'une erreur durant la réplication. On distingue deux types de substitutions: üTransition : substitution d'une purine par une purine ou d'une pyrimidine par une pyrimidine. üTransversion : Substitution d'une purine par une pyrimidine ou inversement.

• Macrolésions : affectent une séquence de bases. On distingue plusieurs catégories: • Réarrangement : la totalité de l'ADN est présente après ce type de mutation: • Inversion : inversion d'une séquence, • Translocation : excision d'un fragment puis sa réintégration dans un autre endroit. üDuplication : un segment d'ADN est présent en double. üDélétion : perte d'un fragment d'ADN. üInsertion : gain d'un fragment d'ADN.

Effets des mutations ponctuelles • Peuvent affecter aussi bien les gènes de structure que ceux de régulation. • A) Mutations "même sens" (same sense) : Elles sont en général détectées uniquement et seulement après séquençage du gène muté. Ceci est dû au fait que la séquence initiale en AA ne change pas (mutation silencieuse). • Exemple: substitution de U par C (au niveau de l'ADN, transition AT → GC) dans le codon GAU ce qui donne GAC. Les deux codons codent pour l'acide aspartique.
 Mutations \"non sens\" (stop) : mutations qui aboutissent à des codons")
• B) Mutations "non sens" (stop) : mutations qui aboutissent à des codons de terminaison (stop) UAA, UAG ou UGA. Ces codons ne sont pas traduits parce que les bactéries ne possèdent pas d'ARNt qui reconnaît ces triplets. Le résultat de ces mutations est l'arrêt de la traduction avant la synthèse complète du polypeptide, ce qui donne naissance à un produit inactif.
: obtenu par substitution d'une paire de bases par")
• Mutations "faux sens" (missense): obtenu par substitution d'une paire de bases par une autre, donc un AA par un autre AA au niveau du polypeptide. • S'il s'agit d'une enzyme: üla protéine peut avoir un site catalytique altéré donc perte totale ou partielle de l'activité enzymatique. üla protéine peut devenir anormalement sensible à un facteur physique ou chimique; ex: mutants thermosensibles. üles unités polypeptidiques peuvent subir une association anormale, ce qui a pour conséquence la perte de l'activité catalytique. üParfois aucun changement n'a lieu; c'est le cas d'une substitution d'un AA par un autre qui a les mêmes propriétés, ex: Glu par Asp. üParfois l'AA n'intervient pas dans les sites actifs et n'est pas déterminant dans la conformation.

• Microdélétions ou microinsertions: La conséquence de telles mutations est le décalage de lecture au niveau de l'ARNm. • Dans ce cas, le polypeptide produit contient la séquence correcte d'AA jusqu'au point où il y a eu mutation à partir duquel la séquence en AA change. Parfois, ce décalage génère un codon non-sens, d'où l'arrêt de la traduction. • Mutations "suppresseurs" (Réversion): Un organisme muté peut subir une deuxième mutation qui ramène au caractère sauvage (initial). Cette seconde mutation est dite "suppresseur" car elle supprime l'effet de la première.
 ou")
4. 4. 1 Caractères de la mutation bactérienne • Elle est spontanée (hasard) ou induite. Elle peut s’établir par l'analyse statistique de la distribution des mutants dans des tubes de bouillon de culture ensemencés en parallèle avec une même suspension microbienne. Peutse fare selon deux tests: • test de fluctuation de Luria et Delbruck, 1943) • test des répliques au tampon de velours, sans contact avec l'agent sélecteur (Lederberg et Lederberg, 1952) • Elle et discontinue (caractère brusque) • Elle est stable: le caractère acquis par la mutation est transmis à la descendance et se maintient. La réversibilité de la mutation ( « reverse mutation » ) est cependant possible

• La mutation est un phénomène rare qui n'affecte qu'une faible fraction de l'ensemble des cellules bactériennes au sein d'une large population. La proportion des mutants que l'on peut observer dans la population bactérienne d'origine dépend de trois paramètres indépendants de : • la probabilité qu'une cellule bactérienne mute dans une unité de temps donné, correspondant à un certain nombre de générations. Cette probabilité s'appelle le taux de mutation ; • la distribution dans le temps des évènements mutationnels durant la période de culture, des mutations très précoces produisant de très larges clones de descendants du mutant) ; • le taux de croissance du mutant comparé à celui du type parental sauvage ( « fitness »

• Elle est indépendante et spécifique: La mutation n'affecte habituellement qu'un seul caractère en respectant les autres (ex. : M. tuberculosis sensible à tous les antibiotiques → M. tuberculosis résistant à la streptomycine et sensible à tous les autres antibiotiques). Mais lorsque les mutations affectent une séquence de gènes fonctionnant ensemble (un opéron), elles peuvent affecter plusieurs caractères (mutation pléiotrope).

4. 4. 2 Les variations génétiques par transfert de matériel génétique • Le transfert vertical se produit lorsque l'organisme reçoit du matériel génétique à partir de son ancêtre. • Le transfert horizontal de gènes, ou transfert latéral de gènes : un organisme intègre du matériel génétique provenant d'un autre organisme sans en être le descendant. • systèmes principaux d'échange de matériels génétiques chez les bactéries: • la conjugaison : transfert d'ADN d'une bactérie à une autre, via un contact de cellule à cellule (par pili sexuel par exemple) ; • la transformation: prise d'un matériel génétique extracellulaire (provenant d'une bactérie morte. . . ) par une autre bactérie. • la transduction : processus par lequel l'ADN d'une bactérie est transporté dans une autre bactérie par le biais d'un virus.

A. Transformation génétique chez les bactéries • La transformation est le transfert passif d'ADN d'une bactérie donatrice à une bactérie réceptrice, dite en état de compétence. • Ce transfert est partiel et limité à quelques espèces bactériennes, • Il entraîne l'acquisition par la bactérie réceptrice de nouveaux caractères génétiques stables et transmissibles.

• Découverte en 1928 par Frederick Griffith qui démontre que l'inoculation sous-cutanée à la souris d'un mélange de pneumocoques capsulés (virulents) tués par la chaleur et de pneumocoques acapsulés (non virulents) vivants, entraîne une septicémie mortelle à pneumocoques capsulés vivants. • Il y a donc eu transformation ou « réversion » des pneumocoques acapsulés (R) en pneumocoques capsulés (S). • La transformation est plus facile lorsque les pneumocoques acapsulés vivants et les pneumocoques capsulés tués sont du même sérotype. • En 1944, Avery Mac Leod et Mc. Carty démontrent que le « principe transformant » est l'ADN bactérien. L'activité transformante est perdue en présence de désoxyribonucléase.
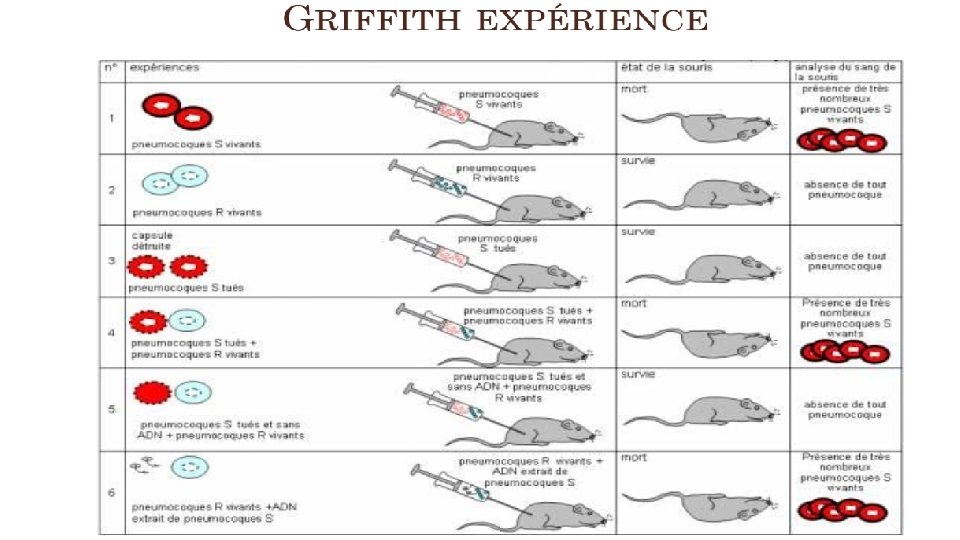

• en injectant à une souris des pneumocoques de souche R ("rough" et non virulents) + une petite quantité de pneumocoques S ("smooth", encapsulés et virulents) tués, la souris meurt et on récupère des pneumocoques deux souches dans son sang. • Si la souche R non virulente dérive d'un pneumocoque de type antigénique III par exemple, et qu'on y ajoute du pneumocoque de type S (virulent) tué de type II, les pneumocoques encapsulés que l'on récupère seront de type antigénique II. • Ainsi les deux caractères d'une souche bactérienne, la virulence et l'antigène de type II ont été transférés à une autre souche. • Conclusion: Les bactéries S mortes ont transmis un facteur transformant les bactéries R en bactéries S.

• La transformation naturelle ou physiologique exige l'état de compétence qui n'apparaît qu'à certains stades de la division cellulaire et seulement chez une fraction de la population bactérienne. • Cette transformation naturelle peut s'observer chez un nombre limité d'espèces bactériennes à Gram positif (Streptococcus et Bacillus) ou à Gram négatif (Neisseria, Branhamella, Acinetobacter, Haemophilus) • Elle se produit en plusieurs phases: • Apparition de l'état de compétence, fixation puis pénétration et intégration de l'ADN donneur dans le génome de la bactérie réceptrice. Chez les bactéries à Gram positif, intervient un activateur spécifique d'espèce, excrété par la bactérie et qui se fixe à la surface de la bactérie. Il y a ensuite synthèse d'une protéine fixatrice de l'ADN, d'une autolysine et une endonucléase. L'ADN fixé est ensuite partiellement hydrolysé puis converti en un fragment monocaténaire.

• Chez les bactéries à Gram négatif, l'état de compétence est aussi en relation avec la synthèse d'un activateur de paroi qui est excrêté par la bactérie à la phase exponentielle de croissance (H. influenzae) ou à la phase stationnaire (Acinetobacter). • La fixation de l'ADN donneur sur la paroi se fait au niveau de sites récepteurs, dans des conditions strictes de métabolisme cellulaire, de p. H, de température et d'osmolarité.

B. La conjugaison • Transfert d'ADN entre une bactérie donatrice et une bactérie réceptrice à travers un contact et l'appariement entre les bactéries • Dans la bactérie donatrice ou mâle existe un facteur de sexualité ou de fertilité (facteur F). Celui-ci permet la synthèse de pili sexuels et donne la polarité au chromosome. • Le transfert d'ADN chromosomique est à sens unique, orienté, progressif et quelquefois total. • Mise en évidence grâce à l'expérience de Lederberg et Tatum (1946).

Mécanisme de conjugaison Formation couple de conjugaison : gènes tra Transfert de l’ADN : gènes mob GEF L 2 S 4 2008 -2009
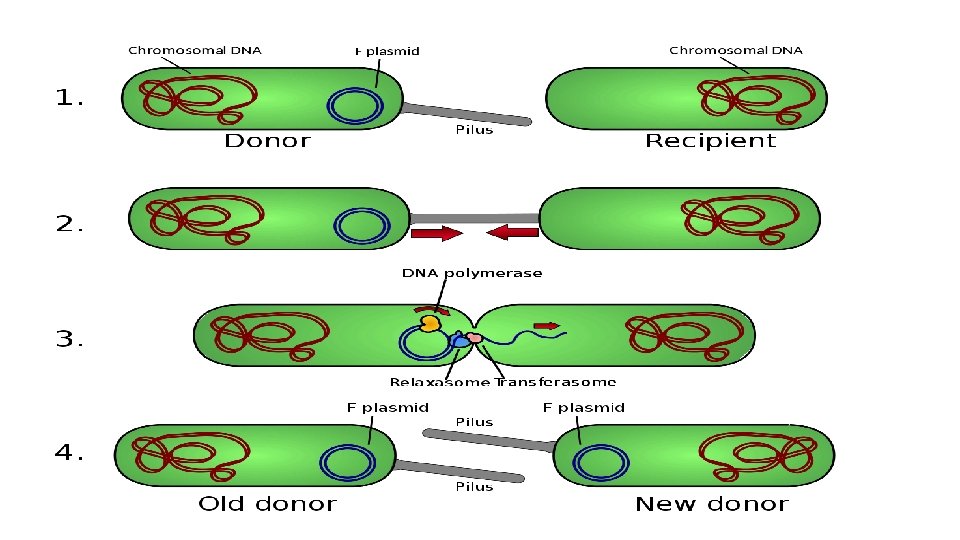

• L'expérience de Lederberg et Tatum • Dans un milieu de culture liquide, on mélange deux types de mutants auxotrophes d'E. coli, • Des mutants exigeants seulement en thréonine (T-) et en leucine (L-) e • Des mutants exigeants seulement en méthionine (M-) et biotine (B-). • Après plusieurs heures de contact entre les mutants TL- M+ B+ et les mutants T+ L+ M- B-, on isole des E. coli T+ L+ M+ B+

La conjugaison

Caractères de la conjugaison • Spécifique: ne se produit qu'entre bactéries d'une même espèce (spécificité), et surtout chez les bactéries à Gram négatif telles que les entérobactéries (E. coli, Salmonella. . . et Pseudomonas aeruginosa). • Différentiation sexuelle: transfert est à sens unique (bactérie donatrice-bactérie réceptrice) repose sur la présence chez la bactérie donatrice du facteur sexuel ou facteur de fertilité (F) à laquelle il confère la polarité ou le caractère mâle (F+). Le facteur sexuel est le premier plasmide connu. L'information génétique qu'il porte code pour la biosynthèse de pili sexuels, pour son insertion possible dans le chromosome bactérien et pour la mobilisation (le transfert) de ce dernier vers des bactéries réceptrices (F-).

• Contact ou appariement: Le transfert chromosomique n'est possible qu'après appariement par couple des bactéries donatrice et réceptrice. • Transfert de l'ADN: Le transfert ne commence quand le pont cytoplasmique formé. • Le transfert de gènes par conjugaison est un facteur majeur d'évolution du patrimoine génétique bactérien, qui joue un rôle essentiel en bactériologie médicale (résistance aux antibiotiques. . . ).

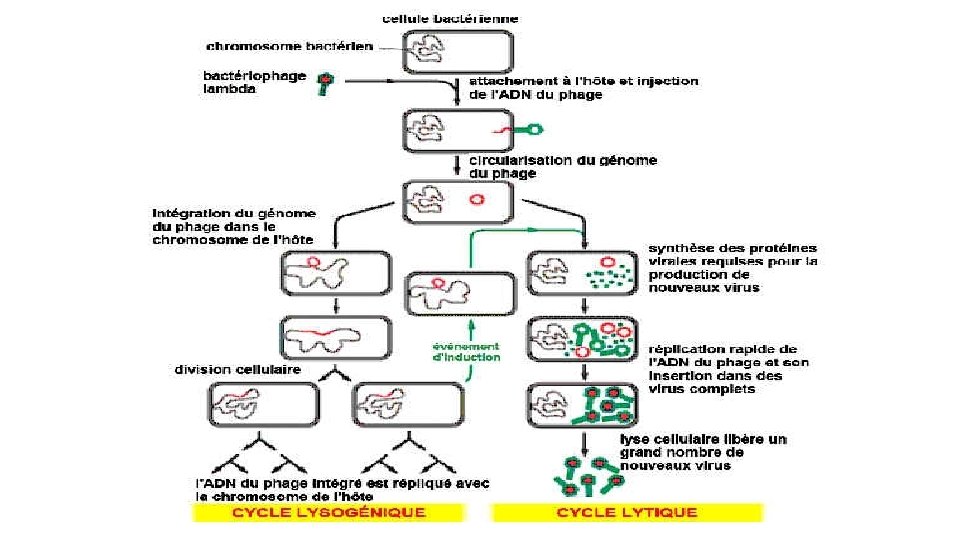
C. Transduction • La transduction est le tranfert d'ADN bactérien par l'intermédiaire de bactériophages (ou phages) ou virus des bactéries • Ces virus existent sous la forme: virulente ou tempérée. • Les phages virulents se multiplient dans la bactérie (ou mieux sont répliqués par la bactérie) et la lysent. • Les phages tempérés s'intègrent dans le chromosome bactérien sans induire la réplication et sont répliqués en même temps que lui. Le bactériophage est alors appelé prophage et la bactérie porteuse est une bactérie lysogène.

Caractères de la transduction • Incidence: la transduction est liée à l'existence de bactéries lysogènes, à Gram positif (staphylocoque, streptocoque, Bacillus ou à Gram négatif (entérobactéries, Pseudomonas). • Type de transduction: • la transduction est complète ou généralisée lorsque les gènes transférés s'intègrent dans le chromosome de la bactérie réceptrice et que celle-ci les transmet à sa descendance. • la transduction est abortive si les gènes transférés ne sont pas intégrés dans le chromosome, ce qui est fréquent, Dans ce cas, les gènes passent de la cellule mère à une seule cellule fille, etc. . . Il n'y a pas généralisation du caractère transféré à l'ensemble descendants. • Conversion lysogénique. Le génome du bactériophage apporte par luimême un nouveau caractère très important pour la bactérie réceptrice. Exemple, la sécrétion de la toxine diphtérique, la sécrétion de la toxine érythogène du streptocoque A (scarlatine) ou la présence de certains facteurs antigéniques. On dit alors qu'il y a eu conversion lysogénique.

Rôle des plasmides • Les plasmides sont des molécules d'ADN bicaténaire, circulaires et cytoplasmiques, de petite taille (5 à 4000 fois plus petit que le chromosome), se replicant d'une manière autonome et non indispensables au métabolisme normal de cellule-hôte. • Leur transmission d'une cellule bactérienne à une autre peut s'effectuer par conjugaison (Tra+) ou transduction. • Leurs gènes peuvent coder pour la synthèse de protéines qui confèrent des propriétés biologiques diverses à la bactérie : • résistance aux antibiotiques (bêta-lactamines, aminosides, phénicols, cyclines, macrolides) chez les bactéries à Gram positif ou négatif ; • résistance aux antiseptiques mercuriels, aux métaux lourds (antimoine, argent, bismuth. . . ) ; • résistance aux bactériophages. • Adaptation à un environnement hostile.

• Le plasmide peut médier la virulence d’une bactérie? C’est le cas du pouvoir pathogène des colibacilles entéropathogènes (diarrhées des voyageurs), pouvoir pathogène des staphylocoques dans l'impétigo (exfoliatine). • Les plasmides peuvent également coder pour la synthèse de bactériocines qui inhibent la croissance d'autres bactéries (ex. : colicines létales pour les entérobactéries). • Ils peuvent aussi porter les gènes qui codent pour le métabolisme du lactose ou de la lysine chez les Proteus, la production de H 2 S chez E. coli, la dégradation du toluène ou de l'octane chez les Pseudomonas. . . • Les plasmides possèdent des gènes qui assurent leur réplication autonome.

• La transposition: intégration directe d'une séquence de gènes, de taille limitée, au sein d'un génome (chromosomique ou plasmidique), en l'absence d'homologie de séquence nucléotique (recombinaison illégitime). • Le transposition est un mécanisme d'adaptation génétique particulièrement efficace des bactéries à leur environnement. • Les transposons (Tn) sont des gènes qui et qui portent les déterminants de la transposition (excision, intégration, transposition) et des gènes qui codent pour d'autres fonctions, par exemple la résistance aux antibiotiques. • La majorité des transposons identifiés proviennent des plasmides de bacilles à Gram négatif, mais certains proviennent de cocci à Gram positif comme le transposon de résistance à l'érythromycine chez Staphylococcus aureus.

• En conclusions, • La conversion et la transduction sont des phénomènes qui font intervenir un bactériophage. • Dans la conversion, c'est le génome du bactériophage qui est responsable du nouveau caractère acquis par la bactérie ; • Dans la transduction, le bactériophage a seulement un rôle de vecteur et le génome transféré provient d'une autre bactérie. .

4. 5 Les bactériophages • Virus n’infectant que des bactéries. • parasites intracellulaires obligatoire, • se multiplient à l'intérieur des bactéries en utilisant tout ou partie des mécanismes de biosynthèse de l'hôte • Chaque Bactéries possèdent son propre phage. • Ils sont facilement isolables car ils se développent parallèlement aux Bactéries. • Ils se retrouvent dans les déchets, sols, excréments des Hommes et animaux et même dans des zones extrêmes (dorsales océaniques, sources thermiques, . . . ).

• les bactériophages sont très diversifiés tant au niveau de leur morphologie que de la composition de leurs génomes. • Ils représentent le plus important réservoir de matériel génétique encore inconnu • Chaque Bactéries possèdent son propre phage. • Ils sont facilement isolables car ils se développent parallèlement aux Bactéries • Très utilisés dans la recherche en génétique moléculaire

4. 5. 1 Morphologie et structure • un phage possède du matériel génétique encapsidé dans une structure protéique complexe constituée le plus souvent d'une tête et d'une queue. • La capside du phage a pour rôle primaire de contenir et de protéger le génome viral. Sa géométrie peut être très variable. • Ne procèdent qu'un seul type d'acide nucléique : ARN ou ADN. • Se produit à partir de son seul acide nucléique. • N'a aucune information génétique concernant les enzymes du métabolisme intermédiaire (susceptibles de produire de l'énergie). • La multiplication des virions implique l'utilisation des structures de la cellule hôte, et spécialement de ses ribosomes.

1. tête ; 2. queue ; 3. acide nucléique ; 4. capside ; 5. col ; 6. gaine contractile ; 7. fibres caudales ; 8. spicules ; 9. plaque terminale.

• On peut regrouper les bactériophages en trois grands groupes morphologiques : • cubique. • Filamenteux. • Mixte.

Classification des Phages • Quatre critères essentiels : • La nature des matériels génétiques (ADN ou ARN) ; • le type de symétrie suivant lequel le virus est construit (hélicoïdale, icosaèdrale, combiné) ; • le caractère nu ou enveloppé de la nucléo capside. • les données quantitatives concernant le virion a symétrie icosaèdrale, longueur et épaisseur des nucléo capside pour les virus a symétrie hélicoïdale.

1. Classification des Phages. • Basée sur la structure de la particule virale et sur la structure du génome. Il existe 4 types: • A) Phage de la série T • Ces phages sont très utilisés dans les manipulations de Biologie moléculaire, en particulier les • l'ADN est le support de l'information génétique. • Les T 7 sont utilisés pour des vecteurs de clonage comme promoteur fort. • dans leur génome, à la palce de la Cytosine il y a de l'Hydroxy. Méthyl. Cytosine. Il leur permet de résister aux enzymes de restriction lors de l'infection. • Tous les phages de la série T possèdent un ADN linéaire bicaténaire et possèdent une structure avec « tête et queue » .
 Phage Tempéré • Ils présentent deux cycles de multiplication, contrairement aux")
• b) Phage Tempéré • Ils présentent deux cycles de multiplication, contrairement aux phages T. • un cycle lytique, comme les phage T, • un cycle lysogène: le phage s'intègre dans le génome bactérien au lieu de rester virion. • C) Petit phage à ADN • Ce sont des phages de petite taille avec deux sortes de nucléocapside. • nucléocapside icosaédrique (comme X 174, S 13). L'étude de ces phages a été à l'origine de la découverte des gènes superposés. • nucléocapside filamenteuse (comme phage fd, phage M 13). Ces phages sont très utilisés car ADN simple brin (donc facilement séquençable).
 Phage à ARN: • Ils appartiennent à la famille des Leviridae")
• d) Phage à ARN: • Ils appartiennent à la famille des Leviridae comme le phage Qß, Ms 2. Ils servent de modèles pour la traduction de l'ARN. • Les phages présentent différents types de génomes: ADN double-brins, ADN simple-brin, ARN; et différents types de structures de nucléocapside: filamenteuse, isocaïdrique ou comme les T 2.

Reproduction : cycles lytique et lysogénique • injectent leur matériel génétique dans des bactéries hôtes. • Utilise les enzymes et ribosomes de l'hôte (et à certaines protéines virales selon les cas), . Deux situations possibles: • le génome viral peut être répliqué pour former de nombreuses copies du virus qui sont libérés avec la lyse de la bactérie-hôte : on parle de cycle lytique ou cycle de production. • leur matériel génétique peut être répliqué et s'intègre au chromosome de la bactérie (ou existe sous forme de plasmide), mais n'est pas exprimé pour former des virions. Le virus est dit prophage, lequel est transmis à la descendance de la bactérie infectée (lignée lysogénique) et on parle de lysogénie ou de cycle lysogénique. • En réponse à une induction (ex : stress de la bactérie), l'infection lysogénique peut basculer vers un cycle lytique.
- Slides: 48