CARACTERISTICILE GENELOR NEALELE 1 Inlantuirea genica lincaj harti
 § harti genetice § dezechilibrul lincajului 2.")


, inca vedeta a cercetarii genetice*, a reprezentat la inceputul")

. Din")


 recombina nti")

 intre 2 loci conteaza in cartografierea fizica; dar distanta fizica")

 Cea mai mare valoare POSIBILA")


 avand loci")



 este format din gene localizate la nivel 6 p.")

, in haplotipuri alelele apar independent")


 masoara in centimorgani distanta dintre 2 markeri")



 avand gena bolii, se observa inlantuirea")












 mutatii: - O alela noua,")
 este")
 • Asocierile apar cand se amesteca doua sau mai multe")









 si gena distrofiei miotonice (cz.")

")




- Slides: 61
CARACTERISTICILE GENELOR NEALELE 1. Inlantuirea genica (lincaj) § harti genetice § dezechilibrul lincajului 2. Transpozoni
Inlantuirea genica “ Segregarea independenta a caracterelor” “PRINCIPIUL ASORTARII INDEPENDENTE” a genelor nealele (situate pe loci diferiti) este: - totdeauna ADEVARAT pt. gene situate pe cz. diferiti - uneori adevarat pt. gene situate pe acelasi cz. Gene sintenice (= avand locii pe acelasi cromozom) se pot transmite: Ø independent, cand sunt separte prin crossing- over Ø inlantuite (lincate), cand au locii apropiati pe cromozom
Segregarea independenta a caracterelor in F 2 Om de stiinta Calugar augustin
Musculita de otet (Drosophila melanogaster), inca vedeta a cercetarii genetice*, a reprezentat la inceputul secolului 20 obiectul de studiu la Universitatea Columbia (NY). * http: //www. brain. riken. jp/bsi-news/bsinews 30/no 30/networke. html Thomas Hunt Morgan a utilizat D. m. deoarece: - se reproduce prolific - in doar 2 saptamani apare noua generatie - este usor si ieftin de crescut si analizat (lupa si/sau microscop) - are doar 8 cromozomi In Theory of the Gene (1926) va recunoaste ca dorea initial sa testeze teoriile lui Mendel, dar rezultatele celor 15 ani de cercetari pe D. m. l-au facut sa confirme teoriile predecesorului. Dupa un an Morgan va observa o D. m. cu ochi albi. Impreuna cu colaboratorul sau Alfred Henry Sturtevant va crea prima harta a unui cromozom. Pentru contributia adusa la fundamentarea geneticii clasice, Morgan a primit Premiul Nobel (1933)
In 1913, in Journal of Experimental Zoology Sturtevant propune ca genele au o pozitie fixa pe cromozom fiind dispuse linear. El a creat astfel prima harta cromozomiala, ceva asemanator imaginii de mai jos.
Cartografierea cromozomiala consta din atribuirea unei gene un loc specific pe cromozom (locus). Din analiza datelor obtinute in urma incrusisarilor experimentale la D. m. s-a obtinut harta cromozomiala. Harta cromozomiala este o diagrama, care prezinta ordinea lineara a genelor pe cromozom. Sturtevant a comparat frecventa crossing- overului pentru numeroase gene. Procentul de astfel de recombinari pentru doua caractere este proportional cu distanta dintre genele care determina acele caractere. Astazi exista noi tehnici utilizate in cartografiere. Alaturat este o harta cromozomiala umana, unde sau investigat lipoproteine si trigliceride.
Segregarea independenta Conform legilor lui Mendel, cele doua caractere codificate de alelele: A, a si respectiv B, b segrega independent (A, B) In gameti sansele de intalnire a genelor este aceeasi (25%). Chiar daca perechile de gene se gasesc pe acelasi cromozom, daca ele sunt la distanta una de alta, atunci sansele ca ele sa se transmita independent sunt aceleasi ca si in cazul genelor situate pe cromozomi diferiti
Prima harta genetica pe baza lincajului Pentru analiza se incruciseaza un heterozigot si un homozigot recesiv (“back-cross”). Se observa numarul mare al descendentilor identici fenotipic cu genitorii si numarul mic al celor care dovedesc crossing-overul in meioza parentala.
Inlantuirea genica (lincajul genic) recombina nti
Conceptul de inlantuire Are la baza posibilitatea ca 2 loci sa se afle pe acelasi cromozom la distanta mica unul de altul a. i. cel mai probabil se transmit impreuna (co-segrega) in timpul meiozei ↓ Recombinarile intre cei 2 loci sunt cu atat mai rare cu cat sunt situate mai apropiate perechile de gene
Distanta fizica (in pb) intre 2 loci conteaza in cartografierea fizica; dar distanta fizica # distanta genetica utilizata in hartile genetice! Nr. mare de recombinari (crossing – over) A A B B Recombinari putine sau absente (valori scazute ale fractiei de recombinare) Harta genetica = o reprezentare a pozitiei relative a genelor pe cz. , bazata pe frecventa/fractia de recombinare meiotica intre 2 segmente de ADN o harta de inlantuire (
Proportia descendentilor recombinanti in raport cu nr. total de descendenti este denumita fractie de recombinare (θ) Non recombinanti In acest caz, cat este θ? Θ= 2/4=> 50%
θ reflecta distanta dintre 2 loci (si nu gene!) Cea mai mare valoare POSIBILA a θ 0, 5, adica 50% NU EXISTA INLANTUIRE corespunde frecventei de segregare a genelor avand loci pe cz. diferiti! Daca θ< 0, 5 EXISTA INLANTUIRE Ex. Daca θ = 0, 05, inseamna ca in medie alelele sintenice vor segrega impreuna de 19 ori din 20 segregari posibile o recombinare (c-o) va avea loc intre ele in medie doar intr-o meioza din 20*
Frecventa de recombinare • θ este deci raportul dintre numarul de descendenti recombinanti si numarul total de descendenti • Unitatea de masura a distantei genetice este denumita Morgan 1 centi. Morgan (c. M) = distanta genetica in care se produc recombinari cu o frecventa de 1% 1 c. M ~ 1. 000 pb (1. 000 kb)
Inlantuirea genica - loci foarte apropiati, inlantuiti, “lincati”, care nu sunt niciodata separati prin crossingover: distanta genetica este de 0 c. M (θminim = 0/totalul descendentilor = 0) fara recombinare - loci neinlantuiti, “nelincati”, mereu separati: distanta genetica este egala cu 50 c. M (adica: fiecare alela a unui locus are o probabilitate de 50% de a fi co-transmisa cu orice alela de pe celalalt locus, iar atunci θmaxim = 0, 5) asortare
FAZE DE INLANTUIRE Intre 2 perechi de gene alele (A/a si B/b) avand loci situati pe acelasi cz. exista 2 posibilitati de dispunere, numite faze de inlantuire: § cuplare sau cis ( A si b, respectiv a si B) § repulsie sau trans (A si B, respectiv a si b) A b a B Sunt aceste gene alele sintenice? . Locii sunt pe acelasi cz, deci sunt sintenice cele doua perechi de gene alele
Pentru a detecta recombinarile dintre loci este necesar ca: 1. Un parinte sa fie heterozigot pentru acei loci (parinte informativ) 2. Faza de inlantuire sa fie cunoscuta In exemplul alaturat mama I 1 este informativa fiind heterozigot pentru cei doi loci marker (1 si 2), cat si pentru locusul bolii. Daca ea ar fi fost homozigot bb, toti descendentii ar fi mostenit o alela b, indiferent de ceea ce mosteneau pe locusul bolii facand imposibila detectarea prezentei sau absentei recombinarii. Daca nu se stia exact faza de cuplare, nu se putea determina care copii sunt recombinanti (II 1, II 3, II 5) si care nu (II 2, II 4, II 6). Retinita pigmentara este o boala degenerativa care conduce la cecitate (orbire) Retinita pigmentara co-segrega cu locusul 1 si 2. Boala se transmite autozomal dominant si doar contributia materna este aratata la descendenti, deoarece aceasta are gena bolii, D. De asemenea pe acelasi cz. II 1 prezinta genele A si B pentru cei doi markeri. Tatal este sanatos si homozigot (dd) pentru RP 9, cat si pentru markeri (AA si BB).
Recombinarea creeaza noi haplotipuri Haplotip: combinatie specifica de loci, unul continand deseori o gena marker pentru o boala ( = markerul se transmite lincat cu gena bolii) Haplotipul nou ab poate fi rezultatul recombinarii sau mutatiei (recombinarea actioneaza mai rapid) A b a B A B a b
HLA sunt antigene umane leucocitare; genele care le codifica sunt CODOMINANTE, deci o persoana are doua haplotipuri fiecare fiind exprimat in fenotip. Deoarece locii sunt situati suficient de aproape unul de altul pe cromozomul 6, intr-o familie intregul haplotip va fi transmis in bloc la descendenti. Un eveniment rar, o recombinare a avut totusi loc in aceasta familie la I 2 in timpul gametogenezi, astfel II 5 prezinta un haplotip recombinant. Intre populatii exista diferente in ceea ce priveste frecventele alelelor si haplotipurilor HLA: intr-o populatie anumite haplotipuri sunt mai frecvente decat este de asteptat, in timp ce altele sunt foarte rare sau chiar inexistente. Majoritatea combinatiilor alelice, care pot aparea teoretic pentru a realiza un haplotip nu au fost niciodata observate in populatii. Aceasta restrictie in ceea ce priveste diversitatea haplotipurilor posibile intr-o populatie apare datorita unui dezechilibru al inlantuirii
Complexul major de histocompatibiltate (MHC) este format din gene localizate la nivel 6 p. Acestea sunt clasificate pe criterii structurale si functionale in 3 clase. Clasa I si II corespund cu genele sistemului HLA. Acestea codifica proteine cu rol in initierea raspunsului imun. Pentru transplantul de maduva osoasa si de organe este necesara compatibilitatea HLA si AB 0. kb
Dezechilibrul de inlantuire Frecventa teoretica cu care se gasesc anumite haplotipuri intr-o populatie se calculeaza prin produsul frecventelor individuale genelor alele ce il alcatuiesc. Daca se compara frecv. teoretica cu cea reala (din acea populatie) se pot gasi 2 situatii: 1. frecv. reala nu difera semnificativ de cea teoretica (genele haplotipului sunt in echilibru de inlantuire) 2. frecv. reala > frecv. teoretica ( asociere alelica preferentiala, dezechilibru de inlantuire)
Echilibru de linkage In conditii de echilibru (recombinari multiple), in haplotipuri alelele apar independent una de alta. Examplu: Daca alela A are o frecventa de 0, 3 si alele b o frecventa de 0, 4 → frecventa haplotipului Ab: 0, 3 x 0, 4 = 0, 12. .
Cum aflam daca 2 gene sunt situate pe acelasi cromozom? Cromozomi diferiti θ = 0. 5 Acelasi cromozom θ ≤ 0. 5* Cu cat θ este mai mic ca valoare decat 0, 5 cu atat distanta genetica dintre cele 2 gene este mai mica. * egal cand genele sunt pe acelasi cz, dar nu sunt inlantuite
A. Segregare independenta B. Inlantuire C. Dezechilibrul inlantuirii
Harti ale genomului Genetic maps (hartile genetice) masoara in centimorgani distanta dintre 2 markeri situati pe acelasi cromozom. Physical maps (hartile fizice) masoara in perechi de baze distanta fizica dintre cei 2 markeri (secventierea intregului genom: ~3, 3 miliarde pb).
In imaginea alaturata: - prima este harta fizica - in rosu este harta genetica la femei, iar in albastru cea de la barbat
Analiza asocierii si dezechilibrul inlantuirii • Atunci cand intr-o familie o alela marker segrega cu un FENOTIP ANORMAL (boala) inseamna ca exista inlantuire (linkage) intre markerul respectiv si locusul genei bolii. • Ex. In acest arbore genealogic alela marker A este dominanta • Ex. Alela marker A segrega cu boala
Daca in imagine se adauga cromozomii (barele verticale) avand gena bolii, se observa inlantuirea fizica intre locusul marker si cel al bolii (D); respectiv alela A marker segrega cu boala in aceasta familie, deoarece locusul sau este suficient de apropiat de locusul alelei ce provoaca boala, a. i. nu a mai avut loc nici o recombinare (c-o) Modelul de transmitere a markerului este autozomal dominant
Lincaj si recombinare • Daca consideram 2 loci (nu obligatoriu unul al alelei bolii si celalalt marker) ei sunt: § Lincati / inlantuiti daca proportia de gameti recombinanti este <½ § Nelincati / neinlantuit atunci cand fiecare tip de gamet se formeaza cu aceeasi probabilitate (frecvente egale de aparitie in gameti a combinatiilor posibile in caz de dihibridare respectiv: 25% AB, 25% ab, 25% Ab, 25% a. B)
Lincaj si recombinare
Caracteristicile inlantuirii genice • Inlantuirea este o proprietate a doi loci. • Inlantuirea nu variaza intre populatii • Inlantuirea nu depinde de frecventele alelice, dar capacitatea detecta prezenta inlantuirii depinde de acestea. • Inlantuirea necesita observarea transmiterii gametilor de la parinti la descendenti (imaginea precedenta)
Asocierea alelica
Asocierea alelica • Asocierea se refera la populatia formata din gameti si prin urmare nu necesita observarea transmiterii de la generatia parentala la cea filiala. • Doua alele sunt asociate, daca evenimentul ca un gamet sa poarte alela A nu este independent de evenimentul ca acel gamet sa contina alela B. • Adica, cu alte cuvinte, A si B se vor intalni in acelasi gamet ori mai frecvent (=asociere alelica prezenta) ori mai putin frecvent (=asociere alelica absenta) decat este asteptat pe baza de sansa. • Asocierea alelica este utilizata in genetica populatiei.
Asocierea alelica • In ex. anterior, in prima populatie de gameti genele A si B apar exclusiv impreuna, adica sunt total, complet asociate. • In a doua populatie de gameti nu apare o asociere, caci alelele se cupleaza intre ele la intamplare: A si B se amesteca cu a si b. • Este important de retinut ca spre deosebire de inlantuire, asocierea este populational-specifica. Ea depinde de frecventele alelice si de dinamica din interiorul populatiei studiate, adica este diferita de la o populatie la alta.
Lincaj fara asociere: un anumit marker alelic va segrega cu boala, dar alela specifica poate sa apara in diferite – alte - combinatii in alte familiile analizate
In acest caz o alela marker A este cuplata cu alela bolii la un fondator si este transmisa in prima familie pe cz. ‘bolii’; dar deoarece nu exista nici o asociere, cuplarea alelei respective cu cea a bolii este intamplatoare (a doua familie)
Poate varia de la o familie la alta, care alela marker segrega cu boala; in exemplele urmatoare alela A apoi a segrega cu fenotipul bolii
DEZECHILIBRUL LINCAJULUI Dar oare ce se intampla atunci cand exista inlantuire si asociere alelica!?
DEZECHILIBRUL LINCAJULUI • Alela A segrega cu fenotipul bolii in ambele pedigree-uri, adica o alela particulara este cuplata cu alela bolii la fondatori si transmisa ca urmare a inlantuirii. • Datorita asocierii alelice din cadrul populatiei exista tendinta ca aceeasi alela marker sa fie cuplata cu alela bolii in ambele familii studiate. • Deci dezechilibrul lincajului apare atunci cand alelele sunt mai frecvent impreuna decat s-ar datora sansei/intamplarii, ele fiind fizic apropiate pe catena ADN-ului. Multipli arbori genealogici demonstreaza excesul transmiterii aceleasi alele la indivizii afectati. Lincajul si asocierea sunt proprietati genetice diferite
CAUZELE ASOCIERII ALELICE § MUTATIA §MIGRATIA §EFECTUL DE FONDATOR §DRIFTUL GENETIC §SELECTIA
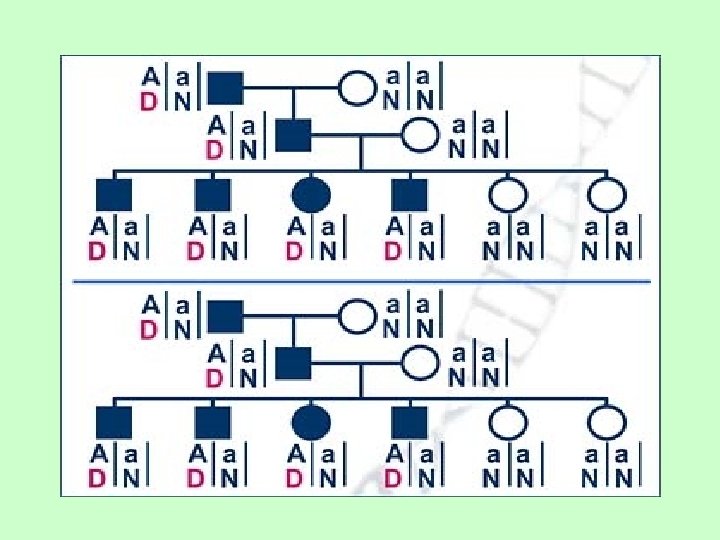
MUTATIA Cel mai cunoscut este modelul noii (de novo) mutatii: - O alela noua, aparuta pe un singur cz. ancestral poate conduce la asocieri intre ea si alele de pe diversi loci din genom - initial exista un marker polimorf (alelele A si a) si alela normala, salbatica (N); apoi apare noua mutatie pe un singur cz. si creeaza asocierea intre alelele A si D (alela mutanta ce provoaca boala)
Boli cauzate de mutatii produse intr-o singura gena • 1. Mucoviscidoza (fibroza chistica) este o b. autozomal recesiva cauzata de mutatii la nivelul genei CFTR*. 7 q 31 Simptomul major este mucusul cu vascozitate crescuta din plamani, ce conduce la infectii. • 2. Anemia falciforma este o b. autoz. rec. cauzata de o mutatie punctiforma in gena ß-globinei. Hematiile in forma de secera impiedica microcirculatia normala si priveaza organele de O 2. • 3. Coreea Huntington o b. autoz. dom. cauzata de mutatii ale genei de la 4 p 16. 3. Simptomele incep la varsta adulta si constau din tulburari psihice si neurologice, cu miscari necontrolate si dementa progresiva. *CFTR= cystic fibrosis transmembrane conduction regulator gene
MIGRATIA (INTRARI SI IESIRI) • Asocierile apar cand se amesteca doua sau mai multe populatii. • Frecventele alelice vor fi diferite in populatiile care se amesteca. Frecv. alelica/genica = frecv. cu care o alela este prezenta pe un locus genic dat intr-o populatie • Pt. a gasi gene ale unor boli, studiind populatiile noi, ce apar in urma acestor migrari, este foarte util cand populatiile initiale aveau boli a caror prevalenta era diferita Prevalenta = nr. de bolnavi dintr-o populatie intr-un anumit moment temporal
Efectul de fondator Initial exista o populatie ‘ancestrala’, cu multi indivizi, unde nu exista asociere intre alela marker si cea a bolii. Apoi un grup de indivizi se desprinde, acestia avand rolul de fondatori: ei poarta numai o parte din totalul de cromozomi ai populatiei ”mama” (genofondul initial). Pe baza de sansa, pot sa apara asocieri!
Ex. pt efectul fondatorului In Vechiul Ordin AMISH din Pennsylvania* apare frecvent displazia condroectodermala, a carei simptomatologie include si polidactilia. Fie mutarea unei parti a populatiei in alta zona, fie reducerea indivizilor ca urmare a schimbarilor de mediu, determina o frecventa disproportionata a genelor fondatorilor in noua populatie. * Bolile transmise genetic sunt mai frecvente in populatiile inchise, casatoriile consanguine previn variatia genetica, fiind pastrate caracterele populatiei initiale. Bolile recesive sunt mai frecvente decat in alte populatii.
DRIFTUL GENETIC Se refera la schimbarile de la parinti la descendenti a frecventelor genice; aceste schimbari apar in populatiile mici prin imperecherea intamplatoare a gametilor, ceea ce conduce la asocieri alelice pe baza de sansa. O frecventa alelica poate creste sau diminua pe baza de sansa; nu fiecare individ va avea urmasi si nici nr. acestora nu este acelasi la diferiti genitori, apoi doar ½ din gene sunt transmise. Efectul, numit drift genetic intamplator, este f. intens: 1. In populatiile mici (100 de indivizi reproducatori sau mai putini) 2. Cand gena este neutra (nici avantajoasa, nici dezavantajoasa)
DRIFTUL GENETIC • Eventual intreaga populatie poate deveni homozigota pt. o alela sau - la fel de probabil – alela poate sa dispara! • Inainte ca aceste lucruri sa se intample, alela reprezinta un polimorfism (poli = multe; morph = forma), adica un fragment de ADN, care mai mult de o forma (alele), fiecare dintre acestea aparand cu o frecventa de cel putin 1% • Polimorfismele contribuie la variabilitate: polimorfismele unei gene pot avea/ nu functii diferite • Polimorfismele pot fi avantajoase, neutre sau dezavantajoase
Un ex. de drift genetic sunt gheparzii, care au cel mai redus nivel de variatie printre mamifere: daca de obicei indivizii inruditi au aprox 80% din gene in comun, la gheparzi se ridica la aprox. 99%! Consanguinizarea a dus la o rata scazuta a supravietuirii, la o calitate redusa a spermiilor si o mai mare susceptibilitate la imbolnaviri. Acestea fiind urmari ale lipsei diversitatii genetice.
SELECTIA NATURALA • Se refera la modul in care speciile se adapteaza la mediu • Conduce la schimbari evolutive, atunci cand indivizi cu anumite caractere au o rata mai inalta de supravietuire sau reproducere decat alti indivizi din cadrul aceleasi populatii; astfel primii vor transmite caracterele genetice ereditare descendentilor • Apare atunci cand combinatiile alelice ofera avantaj (sau dezavantaj) selectiv, conducand la asocieri intre alele. • Realizeaza o diferenta semnificativa dpdv al supravietuirii si reproducerii intre diferitele genotipuri sau chiar intre diferitele gene
Stabilirea relatiei de inlantuire • Este dificila prin studii traditionale bazate pe analiza pedigree-ului – nr. mic de descendenti Daca 2 gene nealele sunt lincate, una dintre ele poate fi folosita ca marker , adica se studiaza co-transmiterea markerului si afectiunii in familiile cu indivizi afectati/neafectati • Studiile populationale pot fi utilizate pt. stabilirea linkage-ului folosind rezultatele recombinarilor.
Marker genetic • Segment de ADN cu o localizare cromozomiala cunoscuta si a carui transmitere poate fi urmarita • Cand 2 segmente cromozomiale (gene nealele) aflate in vecinatate se transmit impreuna, markerul poate fi utilizat pt. depistarea unei mutatii genice ce provoaca aparitia unei boli • Markeri molecular utilizati in genetica umana: – Restriction fragment length polymorphisms (RFLPs). – Minisatellites : VNTR markers – Microsatellite: CA/GT repeatitii – SNPs: Single nucleotide polymorphism (polimorfisme mononucleotidice)
Importanta • Dezechilibru de linkage permite identificarea unor mutatii genice implicate in etiologia unor boli genetice. Ipoteza: Daca un marker genetic este asociat cu un anumit caracter sau afectiune atunci varianta genetica cauzala se gaseste in apropierea sa.
Stabilirea inlantuirii • Examplu: lincaj intre locusul Secretor (marker) si gena distrofiei miotonice (cz. 19) • Distrofia miotonica (Dm): afectiune autosomal dominanta caracterizata prin atrofie musculara progresiva, chelie frontala (expresivitate variabila)
Se utilizeaza un marker genic pt. a stabili transmiterea unei afectiuni In acest pedigree afectiunea se asociaza cu o alela se/Se? se
III-1 si III-2 au genotipuri informative Heterozigot pt. alela markerului Se/se si bolnav (Dm) Heterozigot pt alela markerului (normal) “Back-cross” Homozigot pt. alela recesiva a markerului (normal) Cu cat cele 2 gene sunt mai apropiate pe cromozom, cu atat valoarea predictiva este mai buna
Dm se + Se 96% cazuri 4 c. M Dm + X se + se Se Dm Se 4 c. M In 96% din cazuri indivizii purtatori ai alelei Se nu vor fi afectati 4% cazuri
Deci ce este dezechilibrul de lincaj ? Inlantuirea – se refera la pozitia relativa a unor loci. Lincajul este cosegregarea unei afectiuni sau caracter cu o anumita regiune genomica in diferite familii (intr-o anumita familie este implicata oricare dintre eventualele ale markerului) Asociere – proprietate a alelelor: o anumita alela a genei sau markerului este gasita totodeauna/ niciodata in asociere cu afectiunea sau caracterul Linkage Disequilibrium (DL)– prezenta linkajului Si asocierii; adica cosegregarea unei anumite alele cu afectiunea intr-un nr. semnificativ de familii Deci, DL este tendinta anumitor alele de pe cei 2 loci sa se gaseasca impreuna mai frecvent decat ar fi de asteptat
Inlantuire - versus - Asociere • Studii pe familii in care afectiunea genetica segrega • Studii pe indivizi afectati si neafectati neinruditi • Adecvata cand exista un nr. relativ mic de gene ce confera o susceptibilitate • Porneste de la premisa ca exista 1 sau un nr. foarte mic de variante genetice ce confera un risc crescut • Aplicabila cand anumite alele acestor gene ofera o susceptibilitate crescuta • Nu este necesara cunoasterea defectului genetic ci doar a tipului de transmitere • Aplicabila cand exista un nr. mai mare de gene implicate, fiecare avand un efect modest asupra fenotipului