BTKLER ARASINDAK UYUUM LKLER Doada kendiliinden yetien veya

BİTKİLER ARASINDAKİ UYUŞUM İLİŞKİLERİ Doğada kendiliğinden yetişen veya yetiştirilen bir bitki, aynı arazide birlikte yetiştiği diğer (karışık ekim) veya daha sonra yetiştirilecek bitkileri etkilemektedir. Bitkiler diğer bitkilerden bazılarını olumsuz, bazılarını ise olumlu yönde etkilerken, bazılarını da göz ardı edilebilecek kadar az etkilemektedir. Bu etki çoğunlukla toprak aracılığıyla olmaktadır. Ekim nöbetleri planlanırken, birlikte yetiştirilecek bitkiler arasındaki ve örtü bitkileri, ön bitkiler ile yeşil gübre bitkilerinden sonra yetiştirilecek bitkiler arasındaki uyuşum durumu dikkate alınmalıdır. Bitkiler arasındaki uyuşum ilişkileri etki durumlarına göre allelopati, ön bitki etkisi, sinerjizm ve toprak yorgunluğu olarak incelenebilir.

Allelopati Bir bitki türünün salgıladığı ve/veya artıklarının mikrobiyolojik parçalanmasıyla oluşan kimyasal bileşiklerin, diğer bitki türlerini etkilemesine Allelopati adı verilmektedir (Rice, 1984; Putman ve Tang, 1986). Bir kültür bitkisinin allelopatik olabilmesi için, (a) diğer bitkilerin büyüme, gelişme ve verimini etkilemesi veya (b) aynı bitkinin monokültür tarımda kendi büyüme ve gelişimini etkilemesi veya (c) toprak yorgunluğuna, besin maddesi ve/veya topraktaki mikroorganizma populasyonunda dengesizliğe neden olması veya (d) farklı uygulamalarla yabancı otları kontrol altına alabilmesi gerekmektedir (Einhellig, 1985; Batish ve ark. , 2001).

Allelopatik etki, terpenoidler, fenoller, alkaloidler, kumarinler, taninler, flavonoidler, steroidler, quininler ve çeşitli uçucu yağlar gibi, ikincil metabolizma ürünü allelokimyasallardan kaynaklanmaktadır (Einhellig ve Leather, 1988). Allelopatik etkiye sahip bu kimyasalların bitkilerin doğrudan salgıladığı (Overland, 1966), bitki kalıntılarının ayrışması sonucu (Hedge ve Miller, 1990), bitki yüzeyinden buharlaşma sonucu (Oleszek, 1987), bitki köklerinden salgılanan maddeler (Tang ve Young, 1982) ve bazı bitkilerin polenlerinden (Cruz-Ortega ve ark. , 1988) salgılanan kimyasallar sonucu ortaya çıktığı bilinmektedir (Aslan, 2006).

Doğada, bazı bitki türleri birbirlerinin yaşamlarını desteklerken, bazıları da engellemektedir. Bitkiler yalnızca diğer bitkilerle değil, mikroorganizmalar ve diğer toprak canlıları ile de karşılıklı etkileşim içerisindedir. Yani, her bitki yakın çevresindeki yaşayan tüm organizmalar ile karşılıklı etki-tepki ilişkisi içerisindedir. Bitkiler ve mikroorganizmalar kendilerini ve birbirlerini salgıladıkları bazı maddeler aracılığıyla etkilemektedirler. Bu maddeler ve etkileşim yönü Şekil 6. 1’de görülmektedir.
 Yüksek bitkiler Frenleyici maddeler Mikroorganizmalar Antibiyotikler")
Kolinler (Allelopati) Yüksek bitkiler Frenleyici maddeler Mikroorganizmalar Antibiyotikler

Farklı bitkilerin allelopatik veya autotoksik etkilerine ilişkin çok sayıda araştırma yürütülmüştür. Üzerinde en çok çalışılan bitkilerden birisi de yoncadır. Yonca bazı bitkilerin gelişimini olumlu yönde etkilerken, kendi fidelerine autotoksik etki göstermektedir. İllinois’de yapılan bir araştırma, yaşlı bitkilerin genç fideleri daha çok engellediğini göstermiştir (Çizelge 6. 1). 21 yıl süren bu çalışma, arazide bir yıl yonca olmamasının, yoncanın olumsuz etkilerini gidermek için yeterli olduğunu göstermiştir.


Koca darı hasadından hemen sonra ekilen buğday ve yer fıstığının büyüme ve gelişiminde genelde bir gerileme görülmektedir. Verim azalışı koca darı kök ve anızlarının ayrışması sonucu ortaya çıkan fenolik bileşiklerden kaynaklanmaktadır. Anız toprağa karıştırıldıktan belirli bir süre sonra, fenolik bileşikler parçalanmaya başlamakta ve giderek etkinlikleri azalmaktadır. Toprak işlemesiz tarımda ise, fenolik bileşiklerin etkisi çok daha uzun sürmektedir (Roth ve ark. , 2000; Sene ve ark. , 2001). Arpadan sonra ekilen ekmeklik ve makarnalık buğdayın kök ve sürgün gelişimlerinin, arpanın salgıladığı allelokimyasallardan olumsuz etkilendiği belirlenmiştir (Ben-Hammounda ve ark. , 2001).
")
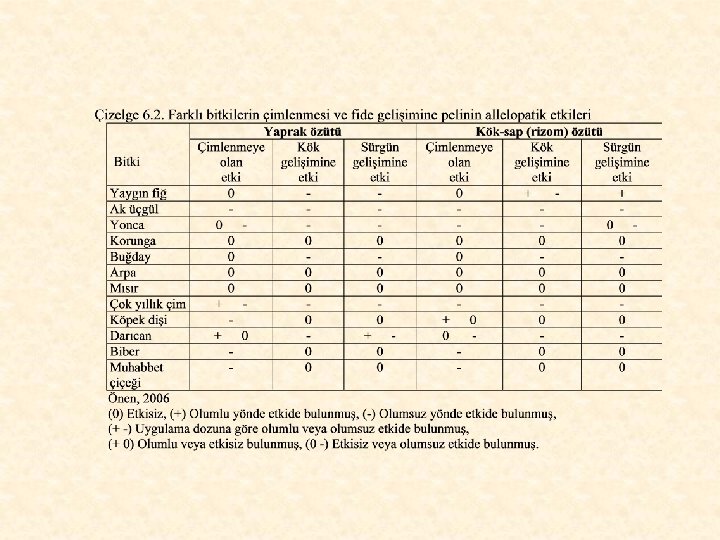
Özellikle Karadeniz Bölgesi’nde sorun oluşturan bir yabancı ot olan pelin (Artemisia vulgaris L. ) allelopatik etkisi çok yüksek bir bitkidir (Önen, 2006). Yapılan araştırmalarda, pelin özütünün soğan, arpa, fasulye, mısır, çok yıllık çim, tilki kuyruğu, imam kavuğu, kısır yabani yulaf, kıvırcık labada ve ak üçgül tohumlarının çimlenmesini olumsuz, nohut ve havuç tohumlarının çimlenmesini ise, olumlu yönde etkilediği belirlenmiştir (Kadıoğlu ve Yanar, 2004; Kadıoğlu ve ark. , 2005). Pelinin allelopatik etkisine ilişkin yürütülen bir başka araştırmadan elde edilen bulgular Çizelge 6. 2’de görülmektedir.

Alelopati ve Yabancı ot Kontrolü Bir ekim nöbeti sisteminde veya birlikte yetiştirme sisteminde yer alan allelopatik bitkiler, herbisit kullanımına gerek kalmadan yabancı otları kontrol etme olanağı sağlayabilirler (Liebman ve Dyck, 1993). Ekim nöbetinde allelopatik bitkilerin kullanımına yönelik araştırmalar yürütülmektedir. Tarla tarımında allelopatik etkisinden yararlanılarak yabancı ot kontrolü amacıyla kullanılabilecek başlıca kültür bitkileri yonca, buğday, koca darı, çeltik, çavdar, ayçiçeği, kanola ve diğer bazı Brassica türleri ile karabuğdaydır (Aslan, 2006).

Brassica türlerinde bulunan bazı allelokimyasallar, özellikle küçük tohumlu yabancı otlara karşı etkili olup, çimlenmelerini önemli ölçüde engellemektedir (Petersen ve ark. , 2001). Kanolanın toprağa karıştırılması ile bazı yabancı otların (sirken, horoz ibiği, darıcan, çoban çantası) çıkışı ve yoğunluğunun, yapay herbisit uygulanmış gibi azaldığı belirlenmiştir (Boydston ve Hang, 1995). Yapılan çeşitli araştırmalarda, kanolanın yabancı ot yoğunluğunu %15 -85 oranında azalttığı saptanmıştır (Grodzinsky, 1992; Boydston ve Hang, 1996; Al-Khatib ve ark. , 1997).

Laboratuvar koşullarında 25 yabancı ot türü ve 32 kültür bitkisine ait tohumların çimlenmesine Antep turpu özütünün etkisi araştırılmıştır. Antep turpu özütünün bulunduğu ortamlarda, içlerinde küsküt, halep otu, tilkikuyruğu ve tarla sarmaşığının bulunduğu 11 yabancı ot türü ile tütün, fasulye, üçgül ve marul tohumlarının çimlenmesinin tümüyle engellendiği belirlenmiştir. Ayrıca, turp özütünün yoğunluğu arttıkça, halep otu köksaplarının (rizom) büyümesi yavaşlamıştır (Uygur ve ark. , 1990). Diğer bazı turp türleri, şalgam ve kanola özütlerinin de, bazı bitkilerin çimlenme ve gelişmesini, artan özüt yoğunluğuna bağlı olarak, azalttığı veya tümüyle engellediği belirlenmiştir (Doğan, 2004; Arslan ve ark. , 2005).
 tohumlarının çimlenmesi üzerine")
Taze yonca sürgünlerinden elde edilen özütün pelin (Artemisia vulgaris L. ) tohumlarının çimlenmesi üzerine etkilerini belirlemek amacıyla yürütülen bir çalışmada, özüt dozu % 25’in üzerine çıktığında çimlenmenin büyük oranda önlendiği saptanmıştır. Aynı amaçla iki yıl yürütülen bir tarla denemesinde, yoncanın etkinliğinin %60 ile %77. 5 arasında değiştiği belirlenmiştir (Önen, 1999). Daha sonra aynı araştırıcı tarafından yapılan saksı denemelerinde, yoncanın toprağa karıştırılan kuru toprak üstü aksamının pelinin gelişimini büyük oranda önlediği belirlenmiştir (Önen, 2006).

Allelopati ve Zararlıların Kontrolü Bitkilerde bulunan allelokimyasallar toprak kaynaklı bitki zararlılarını kontrol etmek amacıyla kullanılabilirler. Karabuğday tarımından sonra, toprakta tel kurdu larva gelişiminin önemli düzeyde engellendiği ve sonuçta tel kurdu populasyonunun azaldığı belirlenmiştir (Valenzuela ve Smith, 2002). Yaprak biti yoğunluğunun fazla olduğu alanlarda, tramin veya hydroksamik asit yoğunluğu yüksek buğday, arpa, çavdar ve mısır ekim nöbetine alınabilecek en uygun bitkilerdir (Aslan, 2006). Yüksek yoğunlukta tramin veya hydroksamik asit içeren buğday, arpa, çavdar ve mısır çeşitleri, hem yaprak bitlerine karşı direnç gösterirler, hem de doğal herbisit olarak kullanılabilirler (Rizvi ve Rizvi, 1992). Mısır-lahana-çavdar, çeltik-tütünçavdar, çavdar-pamuk, çeltik-çeltik, karpuz-çeltik gibi ekim nöbeti sistemleri, kök ur nematodunu başarılı şekilde kontrol edebilmektedir (Davide ve Zorilla, 1983).

Allelopati ve Hastalıkların Kontrolü İyi planlanmış bir ekim nöbeti uygulaması, hastalık ve zararlıların kontrolüne olanak sağlar. Bitkilerin köklerinden salgıladıkları veya mikrobiyolojik ayrışımları sonucu açığa çıkan allelokimyasallar, özellikle toprak kökenli hastalık etmenlerini kontrol altına alabilme potansiyeline sahiptir (Batish ve ark. , 2001). Tütün-çok yıllık çim- mısır ekim nöbetinde, çok yıllık çimin fungitoksinler üreterek kök çürüklüğü hastalığını azalttığı saptanmıştır (Chou ve Patrick, 1976). Tütün-çavdar-mısır ekim nöbetinde de tütünde görülen kök çürüklüğünün azaldığı belirlenmiştir. Nanenin ekim nöbetine alınması ile, pamukta sorun olan Fusarium solgunluğu en az düzeye inmiştir (Li, 1998).

Brassicaceae familyasında yer alan bitkilerin parçalanması ile ortaya çıkan izotiyosiyanatlar, toprakta bulunan bir çok fungal patojene toksiktirler. In vitro çalışmalarda izotiyosiyanatların Colletotrichum, Rhizoctonia, Fusarium, Phytophora, Pythium ve Gaeumannomyces gibi toprak kaynaklı patojenleri kontrol edebildiği saptanmıştır (Kirkegaard ve ark. , 1996; Bianco ve ark. , 2001).

Allelopati ve Toprak Mikroorganizmaları Toprak miroorganizmaları üzerine yapılan çalışmaların çoğunluğu simbiyotik azot fiksasyonu ve mikorizal ilişkiler üzerinedir. Biyolojik olarak bağlanan azot miktarı toprak p. H’sı, sıcaklık, toprak nemi ve havalanması, toprağın besin elementi içeriği, fiksasyonu gerçekleştiren bakterilerin etkinliği gibi birçok faktöre bağlıdır. Aynı zamanda, azot bağlayan mikroorganizmalar, bitkiler tarafından salgılanan allelokimyasalların da etkisi altındadır. Çeşitli baklagiller üzerinde yapılan çalışmalarda, allelokimyasalların nodülasyon ve kök büyümesi üzerine engelleyici veya teşvik edici etkilerinin bulunduğu tespit edilmiştir.
 çeltik bitkisi ile yaptıkları bir çalışmada, çeltik saplarından izole")
Rice ve ark. , (1981) çeltik bitkisi ile yaptıkları bir çalışmada, çeltik saplarından izole edilen fenolik allelokimyasalların Rhizobium bakterilerinin üç hattının gelişimini durdurduğu belirlemişlerdir. Ayrıca bu maddelerin uygulanmasıyla fasulye bitkisinde nodül sayısının ve leghemoglobin miktarının azaldığını bildirmişlerdir. Acı bakla (Lupinus sp. ) kökleri tarafından salgılanan kimyasalların bazı mantar türlerini teşvik ettiği, buna karşın bazı toprak bakterilerini ise engellediği bilinmektedir (Hoagland ve Williams, 1985).

Buğday kökleri çevresindeki mantar ve bakteriler tarafından fenolik asitler, organik asitler, aldehitler, alkoller, ketonlar ve azot içeren allelokimyasallar sentezlenmektedir. Bu kimyasallardan bazı fenoller ve organik asitler buğday ve mısırın çimlenme, büyüme ve gelişimini olumsuz, azotlu bileşikler ise olumlu etkilemektedir (Ma ve ark. , 1997). Kirkegaard ve ark. (1996), yulaftan sonra ekilen buğday veriminde azalma olduğunu, azalmanın besin maddesi veya su yetersizliğinden değil, yulaf saplarının ayrışımı sonucu ortaya çıkan kimyasalların, topraktaki mikroorganizma populasyonlarını kaynaklanabileceğini bildirmişlerdir. etkilemelerinden

Bitki ve toprak arasında yer alan mikorizalar, agroekosistemlerin işleyişine olumlu veya olumsuz etkide bulunabilir. Uygun mikroorganizmalar seçilerek kültür bitkilerinin ve yabancı otların allelopatik etkileri azaltılabilir. Mikorizal zenginlik ve özel konukçu-mantar bileşenlerinin seçimi topraktan daha iyi yararlanmayı sağlar. Allelokimyasallara karşı dirençli mikorizal mantarların geliştirilmesi mümkündür. Mikorizaların allelokimyasallara karşı tepkilerini belirleyen biyolojik ve ekolojik çalışmalar yapılarak, uygun olanlar toprakta allelokimyasalların etkisini azaltmak amacıyla kullanılabilirler (Perry ve Choquette, 1987).

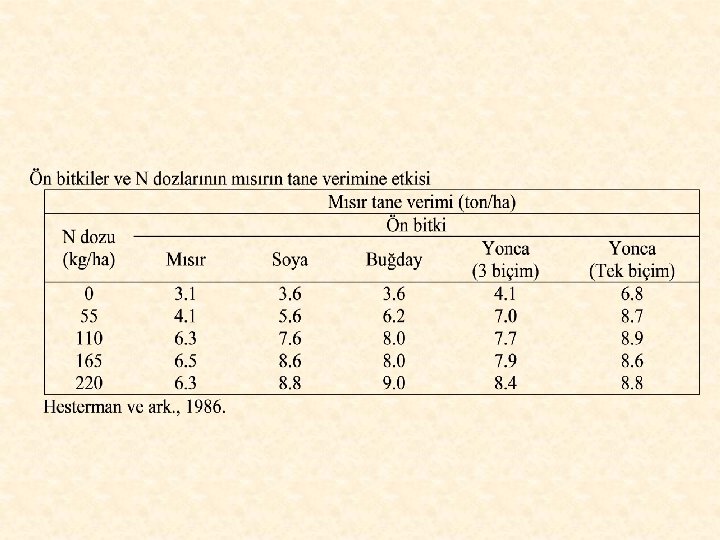
Ön Bitki Etkisi Ekim nöbeti içinde bir diğerinden önce ekilen bitkiye ön bitki, bir diğerinden sonra ekilen bitkiye de izleyen bitki adı verilir. Ön ön bitki veya birinci, ikinci, üçüncü izleyen bitki gibi kavramlar da kullanılır. Birbirini izleyen iki ana bitki ekim nöbeti çifti olarak adlandırılır. Ön bitki etkisi, ön bitkinin kendisini izleyen bitkiler üzerine olan etkisini ifade eder. Ön bitki değeri, değişik ön bitkilerin aynı kültür bitkisi (yani aynı izleyen bitki) üzerinde ölçülebilen etkileri olarak kabul edilir. Herhangi bir bitkinin ön bitki özellikleri, onun yalnız izleyen bitkiler üzerinde ölçülen etkisinin nedenlerini kapsar. Her izleyen bitkinin gerek ekim zamanı ve gerekse diğer yetişme koşulları ile ilgili olarak, belirli ön bitki istekleri bulunmaktadır.

Bitkilerin ön bitki etkisi ve ön bitki değerini bölgenin iklim, toprak vb. çevresel koşullarının etkileyeceği unutulmamalıdır. Bu nedenle, belirli bölgelerde yürütülen ekim nöbeti denemelerinden elde edilen sonuçların, her bölge için olduğu gibi geçerli olacağı düşünülmemelidir. Bir ekim nöbeti sıralamasında yetiştirilen ürün üzerine ön bitkinin çok büyük bir etkisi vardır.
, zamanında tarladan kaldırılmak")
Yonca ve çayır üçgülünün, yağışlı bölgelerde (sulanan yerler de dahil edilebilir), zamanında tarladan kaldırılmak koşuluyla, tahıllar için çok iyi ön bitkiler olabildikleri ortaya çıkmıştır. Ancak, yağışın yetersiz olduğu ve sulanmayan yerlerde, yonca ve çayır üçgülü tahıllar için uygun ön bitkiler değildir. Yonca kendine katlanamayan bitkiler arasında yer alır ve aynı tarlaya ikinci kez yonca ekmek için ekim molası verilmesi gerekmektedir (Sencar ve ark. , 1994; Açıkgöz, 2001).

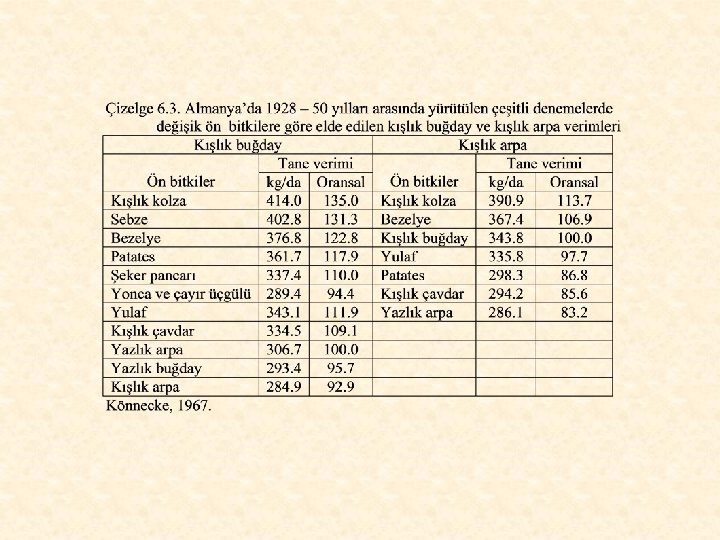
Mısır, soya, kışlık kolza, çeşitli sebzeler ve bezelye kışlık buğday için çok iyi ön bitkilerdir. Patates, şeker pancarı ve yulafın da, kışlık buğday için ön bitki değerleri iyi düzeydedir. Ancak, erkenci patates kullanıldığında, kışlık buğday için patates bezelye kadar, hatta daha da iyi bir ön bitki olmaktadır. Patates gevşek bir toprak yapısı bıraktığından ve özellikle erkenci patatese verilen azotun önemli bir kısmı toprakta kaldığından, arkasından ekilecek buğdaya (veya başka bir tahıla) verilecek N miktarı azaltılmalıdır. Aksi halde, ciddi bir yatma sorunu ile karşılaşılır. Şeker pancarının kışlık tahıllar için ön bitki değerini düşüren en önemli neden, vejetasyon süresinin uzunluğudur. Çok geç hasat edilen şeker pancarından sonra, özellikle kış soğuklarının erken başladığı bölgelerde, toprağın hazırlanarak kışlık tahılın ekilmesi için yeterli zaman kalmamaktadır. Ayrıca, birçok yörede ekim-kasım aylarının genellikle yağışlı olduğu da unutulmamalıdır.

Yulaf kendini takip etmediği sürece ekim nöbetinin her yerine konulabilir. Arpadan en iyi verim nadastan sonra, ardından çapa bitkilerinden sonra, en kötü de kendinden sonra ekildiğinde alınır. Keten anızları arazi üzerinde karın daha çok birikmesini sağladığı için ketenin ardına ekilen arpa yüksek verim sağlar. Keten küçük taneli tahılların ardına ekilmelidir. Eğer ekim geç yapılırsa, ketenin ardından mısır ekilmeli veya arazi nadasa bırakılmalıdır.

Şeker pancarı için en uygun ön bitkiler patates, kışlık çavdar, keten ve mısırdır. Şeker pancarı tarımının yapıldığı yerlerde seçilecek ön bitki ve izleyen bitkilerin, şeker pancarı nematoduna düşman olması gerekir. Çavdar, mısır, yonca, buğday, arpa, patates, fiğ, korunga, nohut ve keten gibi.

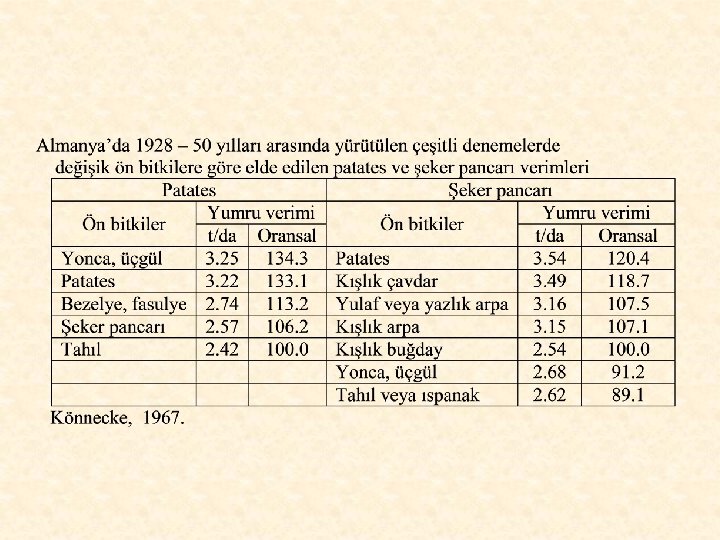
Patates için uygun ön bitkiler yonca, çayır üçgülü, üçgül+çim karışımı, şeker pancarı ve bezelye gibi bitkilerdir. Patates gevşek ve iyi bir toprak yapısı ve bol azot istediğinden, toprağa fazlaca azot ve organik madde kazandıran çok yıllık baklagil yem bitkileri, patates için çok iyi ön bitkilerdir. Yapılan denemelerde, bu bitkilerin patatesin verimini %34 oranında artırdıkları belirlenmiştir. Çok yıllık yem bitkileri toprak yapısını iyileştirdiğinden, keseklenme ve kaymak bağlama önleneceği için, makinalı hasat (söküm) kolaylıkla yapılabilmektedir. Çok yıllık yem bitkilerinin ardına dikilen patates çok güçlü geliştiğinden, virüs hastalıklarına karşı direnci çok artmaktadır. Ancak, çok yıllık yem bitkileri fazlaca su kaldırdığından, yağışların yeterli olmadığı sulanmayan yerlerde erkenci patates türleri yeğlenmelidir.


Ayçiçeği ve aspir geç olgunlaşan derin köklü, alt toprak katını kurutan kurağa dayanıklı bitkilerdir. Ayçiçeği ve aspir küçük taneli tahılların ardına ekilmelidir. Kurak yıllarda, topraktan kaldırılan suyun tekrar yerine gelmesi sınırlıysa, ayçiçeği ve aspir ekimi yerine arazi nadasa bırakılmalıdır. Yemeklik tane baklagiller, ekim nöbetinde herhangi bir çapa bitkisinin yerini alabilirler. Aynı hastalıklar etki ettiğinden, yemeklik tane baklagiller ayçiçeği veya patatesin ardına ekilmemelidir. Soya, mısır veya küçük taneli tahılların ardından ekilmelidir.

İyi planlanmış ekim nöbetlerinde, ön bitkiler izleyen bitkilerin verilen gübreden daha iyi yararlanmasını sağlar. Ekim nöbetinin beklenen diğer yararları da ortaya çıkınca, monokültürlerle karşılaştırılınca, ekim nöbeti sistemi içinde yetiştirilen bitkilerin verimi artar. Yeterli azotlu gübreleme yapılsa bile, ekim nöbeti sistemi içinde yetiştirilen bitkilerin verimleri, monokültürlerden çoğunlukla %10– 40 daha fazla olmaktadır

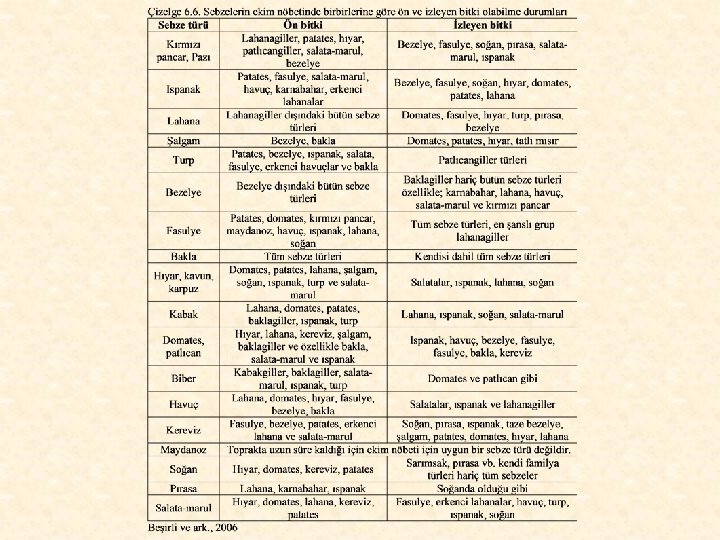
Dünyanın birçok yerinde ve ülkemizde, özellikle küçük ve parçalı arazi yapısına sahip işletmelerde, birkaç bitkinin birlikte yetiştirildiği karışık ekim uygulaması çok yaygındır. Ülkemizin Orta ve Doğu Karadeniz Bölgesi’ndeki köylerde, çiftçiler küçük bahçelerinde mısır, fasulye, patates, kabak veya lahana, bezelye, soğan gibi sebzeleri birlikte yetiştirirler. Ancak, bazı sebzelerin birlikte yetiştirilmesi uygun değildir. Birlikte yetiştirilmeye uygun olan ve olumsuz etki gösteren sebzeler Çizelge 6. 9’da görülmektedir.


Fiğ türleri, mürdümük türleri gibi gövdesi zayıf ve yatma eğiliminde olan yem bitkileri, genellikle bir serin iklim tahılı ile birlikte yetiştirilerek, hem yatma önlenir, hem de yemin protein ve karbonhidrat dengesinin daha iyi olması sağlanır (Acar, 1992). Yonca, çayır üçgülü gibi çok yıllık yem bitkileri fide devresinde yavaş ve zayıf gelişirler. Bu fideleri yabancı ot rekabeti ve diğer olumsuzluklardan korumak ve aynı zamanda ilk yıl daha çok verim almak için, bazen serin iklim tahılları, bezelye, bakla gibi bitkilerle birlikte ekilirler (Önal-Aşçı, 2008). Yine yapay meralar ve yeşil alanlarda uyumlu birkaç bitki birlikte ekilirler. Bunlar da karışık ekim ya da birlikte yetiştirmenin daha farklı uygulanış şekilleridir.

Tarım ve çevre açısından önemli, gerekli olması ve çiftlik gelirlerini artırmasına karşın, ekim nöbetinin bazı istenmeyen yönleri de vardır. Ekim nöbetine katılan yeni bir ürün, yeni ek yatırımları gerektirir. Örneğin, bir çapa bitkisi olan mısır için özel ekim, bakım, hasat makinaları ve ekipmanları gereklidir ve eğer ürün silaj olarak hasat edilecekse çiftlik hayvanlarına da yatırım yapmak gerekir. Yem bitkileri, uygun ekim ve hasat makinaları ve ekipmanları ile çiftlik hayvanları için yatırımı gerektirirler. Şeker pancarı ve patates özel ekipmanlar için ek yatırım gerektiren ürünlere örnektirler. Yeni ürünler ek işgücü gerektirebilir ve yeni pazarlama sorunları yaşanabilir. Bazı durumlarda, çiftlikte ek depolama yapıları gerekebilir.

SİNERJİZM Büyüme etkinliğini artıran bitkiler arasındaki etkileşime sinerjizm adı verilir. Nedeninden tam olarak emin olmamakla birlikte, sinerjizm bitkiler arasında olabileceği gibi, aynı zamanda tek bitkilere özel de olabilir görünmektedir (Anderson, 2005 a).

Bazı bitkiler, özellikle baklagiller, kendisini izleyen bitkiler için sinerjistik olabilir. Örneğin, kışlık buğday, darı veya nadas ile kıyaslanınca, kuru tane için üretilen bezelye kışlık buğdayın su kullanım etkinliğini artırmıştır (Anderson, 2002). Rice (1983), bezelye köklerinin, arpada fotosentezi artıran (stimüle eden) büyümeyi teşvik edici bileşikler salgıladığını bulmuştur.
, fakat etki düzeyi")
Baklagiller bazı tahılların azot kullanım etkinliğini artırabilir (Raun ve Johnson, 1999), fakat etki düzeyi baklagil türleri arasında değişir (Badaruddin ve Meyer, 1994). Hindistan’da, inci darısının (Pennisetum glaucum) azot kullanım etkinliğinin artırılması yönünden, baklagiller arasında benzer bir değişkenliğin olduğu belirlenmiştir (Praveen – Kumar ve ark. , 1997). Buna karşılık, bazen, mısırın ön bitkisi olan soyada görüldüğü gibi, baklagiller azot kullanım etkinliğini artırmazlar (Copeland ve Crookston, 1992). Ürün sıralaması baklagillerin N bağlamasını da etkileyebilir. Tahılların bir yağ bitkisi ve bir baklagille ekim nöbetine girdiği 4 yıllık sıralamanın karşılaştırıldığı bir çalışmada, buğdayla kıyaslanınca kanarya yem kanyaşının ön bitki olduğu durumlarda mercimeğin bağladığı N miktarı % 10 artmıştır (Matus ve ark. , 1997). Kanarya yem kanyaşı mercimekte etkili rhizobium bakterilerine sinerjistik etki yapmış olabilir.

Yakın zamana kadar toprakta alınabilirliği yavaş olan besin elementlerinin alımının, yalnızca bitki kökleri tarafından sağlandığı sanılıyordu. Fakat son yıllarda yapılan bilimsel araştırmalar, bitki besin elementlerinin bitki köklerinin yanı sıra çoğunlukla mikoriza diye adlandırılan ve teşhisi mikroskop altında yapılan, çok miktarda hif üreten mantar türleri tarafından alındığını ortaya koymuştur (Koide, 1991; Marschner, 1995; George ve Marschner, 1996; Ortaş, 1996, 1997). Mikoriza, toprakta varolan sporları aracılığıyla ekosistemdeki bitkilerin yaklaşık %95’inin köklerine enfekte olmaktadır. Mikorizal mantar çok miktarda hif üreterek bitki kök yüzey alanını artırmakta ve kökten çok uzak bölgelerdeki besin elementlerini söz konusu hifleri aracılığıyla alabilmektedir.

Bu işbirliği bitkinin mikorizal fungusa karbon, mikorizal fungusun da bitkiye besin elementi sağlamasıyla gerçekleşmektedir. Etkin bir enfeksiyon gerçekleştiği zaman, mikoriza bitki ile ortak bir yaşam oluşturarak bitkinin su ve bazı mineral besin elementlerini, özellikle de fosfor, çinko ve bakır alımını artırdığı saptanmıştır. Mikoriza enfeksiyonu, bitkilerin azot ve potasyumun yanı sıra demir ve molibden gibi ağır metalleri de daha iyi alabilmesini sağlamaktadır.
’da yürütülen yetiştirme sistemi çalışmalarında, kışlık buğday + nadas sistemi")
1994’den 1999’a kadar Akron (Co)’da yürütülen yetiştirme sistemi çalışmalarında, kışlık buğday + nadas sistemi ile karşılaştırıldığında, kışlık buğday + mısır+nadas sisteminde, kışlık buğdaydan % 10 daha fazla verim alınmıştır. Başlangıçta bu artışın kışlık buğday + mısır + nadas sisteminde, buğdayın daha uzun aralıklarla ekilmesinden dolayı, buğdayda görülen kök hastalıklarının baskılanmasından ileri geldiği düşünülmüştü (Cook ve Veseth, 1991). Oysa, kışlık buğday + nadas ve kışlık buğday + darı + nadas sistemlerinde buğdayın veriminin farklı olmaması şaşırtıcı olmuştur.
 Kullanılan su miktarı (mm)")
Verim (kg/ha) Kullanılan su miktarı (mm)

Bu ekim nöbetlerinde kışlık buğdayın su kullanımı incelendiğinde, alışılmadık bir su kullanım etkinliği eğilimi dikkat çekmiştir. Kışlık buğday + darı + nadas sistemine göre, kışlık buğday + mısır + nadas sisteminde kışlık buğday suyu taneye daha etkin şekilde dönüştürmüştür (Şekil 1). Örneğin, kışlık buğday tarafından kullanılan 300 mm su, kışlık buğday + mısır + nadas sisteminde 3930 kg/ha tane verimine dönüşürken, kışlık buğday + darı + nadas sisteminde bu değer yalnızca 2940 kg/ha olmuştur. Aynı miktarda su kullanıldığında kışlık buğday + darı + nadas sistemine göre kışlık buğday + mısır + nadas sisteminde % 34 daha fazla buğday tane verimi alınmıştır.

Tüm denemelerde yıllık toprak analizlerine göre gerektiği kadar besin maddeleri verilmiş, herhangi bir hastalık salgını da gözlenmemiştir. Ayrıca, kışlık buğday + darı + nadas ile kışlık buğday + nadas sistemlerinin, ikisinde de, kışlık buğdayın su kullanım etkinliğinin benzer olduğu, böylece ekim nöbetinde mısırın varlığının kışlık buğdayın su kullanım etkinliğini artırdığı düşüncesini doğurmuştur.

Bu araştırmalarda mısırın, darının su kullanım etkinliği üzerine de benzer etkiyi yaptığı gözlenmiştir (Anderson, 2004 b). 4 yılın ortalaması olarak kışlık buğday + darı sistemi ile karşılaştırıldığında, kışlık buğday + mısır + darı sisteminde 300 kg/ha daha yüksek darı tane verimi alınmıştır. Başlangıçta bu farklılığı su kullanımının açıklayabileceği düşünülmüştü, ancak her iki ekim nöbeti sisteminde de ekim zamanındaki toprak su düzeyi ve darı tarafından kullanılan su miktarı benzerdi. Kışlık buğday + mısır + darı sisteminde, darı % 24 daha etkili şekilde suyu taneye dönüştürmüştür. Kışlık buğday+darı sisteminde darının su kullanım etkinliği 75 kg ha-1 cm 1 iken, kışlık buğday+mısır + darı sisteminde bu değer 93 kg ha -1 cm -1 ‘ye yükselmiştir.


Daha sonra soya ve mısırda ekim nöbetinin etkilerini tanımlamak için bir dizi araştırma yürütülmüştür. Mısır ve soya ikili ekim nöbetine alındıklarında monokültürleriyle karşılaştırılınca, her ikisinde de % 15 -20 tane verim artışları görülmüştür. Fakat, ekim nöbeti etkisine ürünlerin farklı tip tepkileri verime de yansımıştır (Şekil 2). Mısırdan önce soya ekildiği zaman, sürekli mısır yetiştiriciliği ile karşılaştırılınca mısır daha iri bitkiler oluşturmuş ve tane verim artışı için daha çok su kullanmıştır (Copeland ve ark. 1993; Crokston ve ark. , 1991). Bunun tersine, mısırın ardından soya ekildiğinde, sürekli soya yetiştiriciliğine göre, soya aynı miktarda su kullanmasına karşın, verim % 15 daha fazla olmuş, soyada bitki büyüklüğü değişmemiştir. Böylece, eğer ön bitki mısır ise, ön bitkinin soya olduğu durumla kıyaslanınca, soyanın su kullanım etkinliği artmıştır. Ancak, mısırın su kullanım etkinliği ön bitkiden etkilenmemiştir.
 Kapasite Etkinlik (Soya …>Mısır) (Mısır…> Soya) Sağlanan verim Bitkiler")
Ekim nöbeti etkisi (Su ilişkileri) Kapasite Etkinlik (Soya …>Mısır) (Mısır…> Soya) Sağlanan verim Bitkiler daha çok su kullanır Fakat aynı miktar su kullanımı Şekil 2. Minnesota’ta ekim nöbeti etkisine mısır ve soya tarafından gösterilen farklı tepkiler
, her iki")
Benzer bir eğilim besin maddeleri alımında da görülmüştür. Copeland ve Crookston (1992), her iki bitkide de 10 bitki besin maddesinin biriktirilen miktarı ve yoğunluğunu incelediklerinde, mısır – soya ekim nöbetinde mısırdan fazla verim alındığında daha çok besin maddesi kullandığını belirlemişlerdir. Buna karşılık, monokültür soya ile karşılaştırıldığında, mısırın ardına soya ekildiğinde, aynen suda olduğu gibi, soya besin maddelerini daha etkin kullanmıştır. Böylece, soya ekim nöbetinde aynı miktarda kaynak kullanımı ile daha çok tane üreterek büyüme etkinliğini artırırken, mısırın ekim nöbetine tepkisi artan kaynak kullanımı şeklinde olmuştur. Schneekloth ve ark. (1991), üç sulama rejimi uygulanan ekim nöbetinde mısırın benzer tepkiyi verdiğini, sürekli mısır yetiştiriciliği ve kışlık buğday – mısır – soya ekim nöbetinde mısırın su kullanım etkinliğinin farklı olmadığını bildirmişlerdir.

Akron, CO ve Minnesota’da yapılan çalışmalardan elde edilen sonuçların ışığı altında; mısırın, herhangi bir yolla, kendisinden sonra gelen bazı ürünlerin büyüme etkinliklerini geliştiren bir toprak çevre koşulları bırakabileceğini söyleyebiliriz. Buna karşın, bu yararlı etkileşimin bazı ürünlere özgü olduğu görülmektedir. Mısır soya, kışlık buğday ve darı için uygun ön bitki iken, Schmidt ve Frey (1988), kocadarı için mısırdan daha yararlı ön bitkiler olduğunu, çünkü sürekli kocadarı ve mısır-kocadarı sistemleri ile karşılaştırılınca, yer fıstığı ön bitki olunca kocadarının veriminin %23 arttığını saptamışlardır.

Toprak Yorgunluğu İklim ve diğer çevre koşulları uygun olmasına karşın, topraktan kaynaklanan nedenlerle bitki gelişiminin yavaşlaması veya durmasının nedeni toprak yorgunluğudur. Toprak yorgunluğunun belirtileri daha çok, aynı tarlada aynı bitki türünün her yıl arkaya yetiştirildiği, monokültür tarım uygulanan alanlarda görülmektedir. Toprak yorgunluğunun nedenleri şöyle sıralanabilir: (1) Besin maddesi eksikliği ve toprağın belirli bir mikro besin maddesi yönünden yoksullaşması. (2) Toprakta nematod ve diğer hastalık nedeni zararlıların yoğunluklarının artması. (3) Toprakta belirli mikroorganizma türlerinin aşırı çoğalması ve böylece toprağın biyolojik dengesinin bozulması. (4) Bitkilerin kök, yaprak, çiçek ve meyve gibi organlarından çeşitli toksinler salgılamaları ve bunların toprakta birikmesi, ayrıca bitkilerin toprağa bıraktığı organik maddelerin ayrışması sırasında toksik etkili maddelerin açığa çıkması.

Nedeni ne olursa olsun, uygun bir ekim nöbetinin uygulandığı topraklarda, toprak yorgunluğunun ortaya çıkma riski çok düşüktür. Ekim nöbeti uygulanan topraklarda, besin maddeleri toprak katmanının farklı derinliklerinden dengeli şekilde alınırlar. Her bitkinin organik artıkları veya salgıları ile beslenen mikroorganizmalar farklı olduğundan, herhangi bir mikroorganizma türünün toprakta aşırı çoğalması söz konusu olmaz. Her bitkinin toprağa bıraktığı salgı farklıdır. Ekim nöbeti uygulamalarında, aynı arazide yetiştirilen bitkiler sürekli değiştiğinden, herhangi bir toksik maddenin aşırı birikme şansı yoktur.
- Slides: 56