BOTANYIV ModuleI Embryology BY Dr Sushil Sharma Department

BOTANY-IV Module-I Embryology BY Dr. Sushil Sharma Department of Botany, Govt. College Sangrah, Distt. Sirmour , H. P. India 1
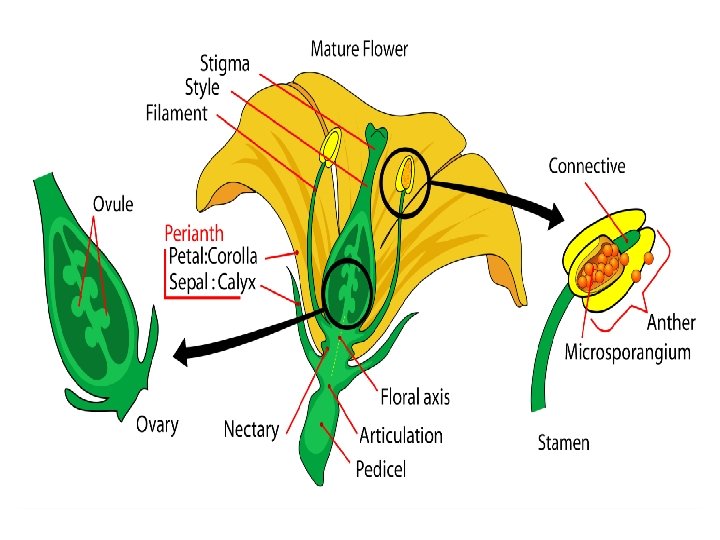


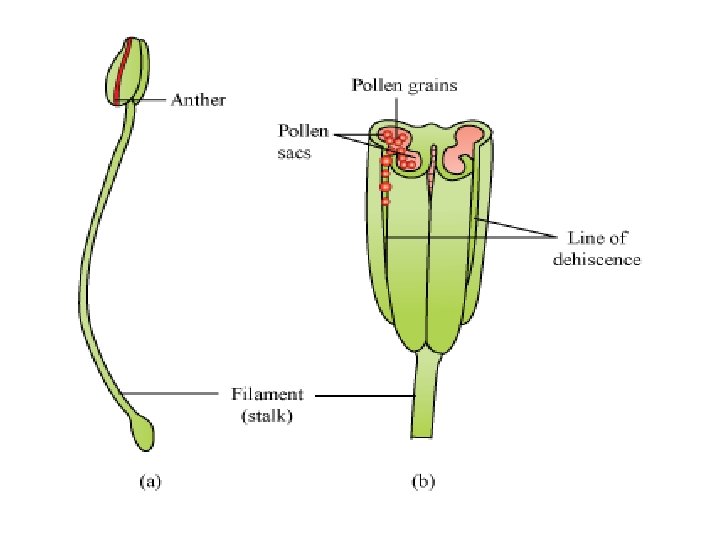
Structure of Anther q The Stamen: • Stamen in a flower consists of two parts, the long narrow stalk like filament and upper broader knob like bi lobed anther. • The proximal end of the filament is attached to the thalamus or petal of the flower. The number and length of stamens vary in different species.

• Most commonly anthers are two lobed and are attached to the filament either at the base or in the middle portion. • The sterile tissue between the lobes is called the connective. • A pollen grain develops from a microspore in the microsporangium and contains the male gametophyte. • The stamens in a flower are collectively called the androecium.

• The androecium in various species of plants form a great variety of patterns, some of them highly complex. • It surrounds the gynoecium and in turn the perianth, if there is one, surrounds the androecium. • A few members of the family Triuridaceae, particularly Lacandonia schismatica are exceptional in that their gynoecia surround their androecia.

Types of Anthers q. The anther can be attached to the filament's connective in two ways: • Basifixed: Anther attached at its base to apex of filament. • Dorsifixed: Anther attached dorsally and medially to apex of filament. • Subbasifixed. Anther attached near its base to apex of filament. • Versatile. Dorsifixed but anther seemingly swinging free on the filament.

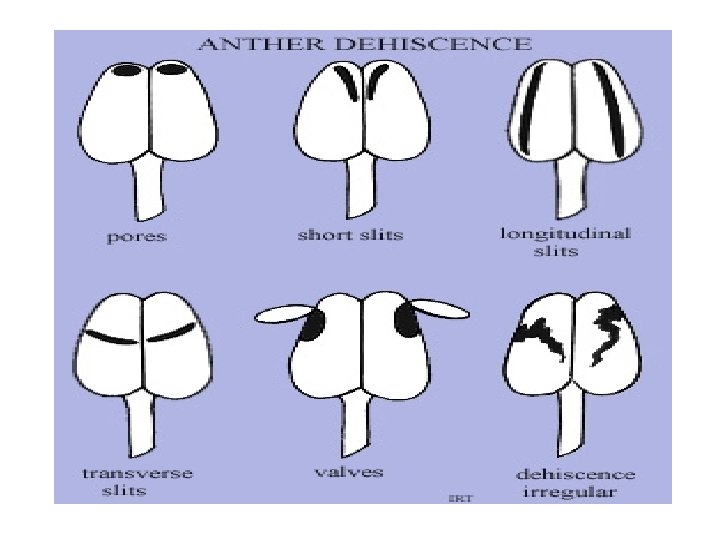
q. Classification based on dehiscence • Extrose: Dehiscing longitudinally outward. • Introse: Dehiscing longitudinally inward. • Latrorse. Dehiscing longitudinally and laterally. • Poricidal: Dehiscing through a pore at apex of theca. • Transverse: Dehiscing at right angles to • long axis of theca. Valvular: Dehiscing through a pore covered by a flap of tissue.

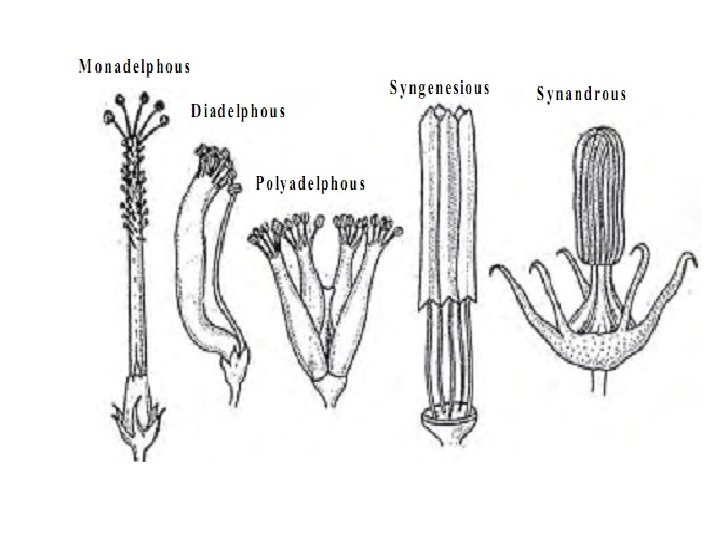
q. Classification based on fusion/grouping: • Monadelphous: Fused into a single, compound structure eg. Family Malvaceae. • Diadelphous: Joined partially into two androecial structures eg. Family Fabaceae. • Pentadelphous: Joined partially into five androecial structures eg. • Polyadelphous: More than five androecial structure eg. Family Bombacaceae


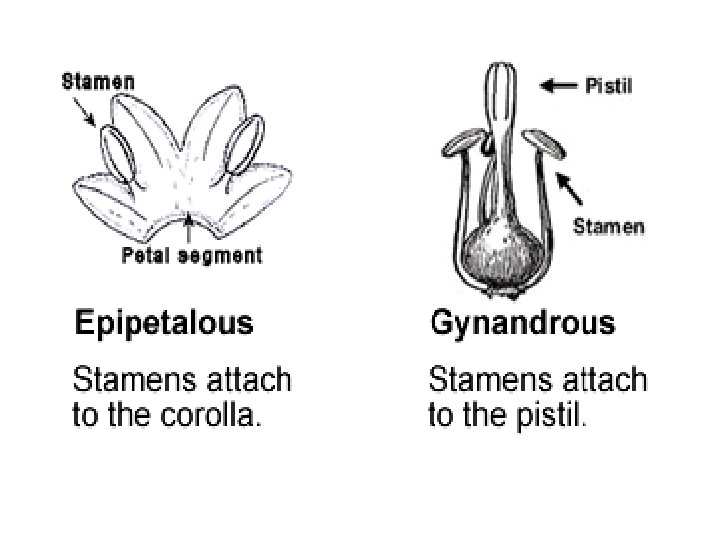
• Synandrous: Only the anthers and stamens are connate (such as in the Asteraceae). The fused stamens are referred to as a synandrium. • Syngenesious: Stamens fused by anthers. • Epipetalous: Adnate to the corolla • Epiphyllous: Adnate to undifferentiated tepals (as in many Liliaceae)

q. They can be different lengths: • Didymous: Two equal pairs • Didynamous: A set of four Occurring in two unequal pairs of different length. Eg. Family Lamiaceae • Tetradynamous: Occurring as a set of six filaments with four long and two shorter ones. Eg. Family Cruciferae

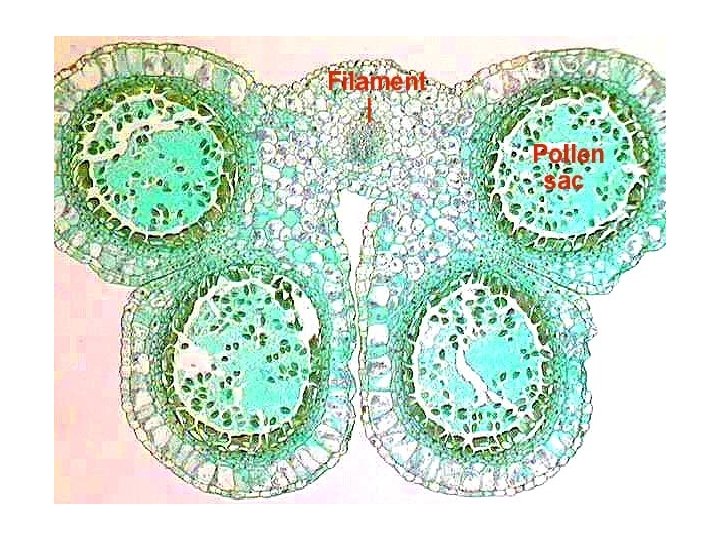
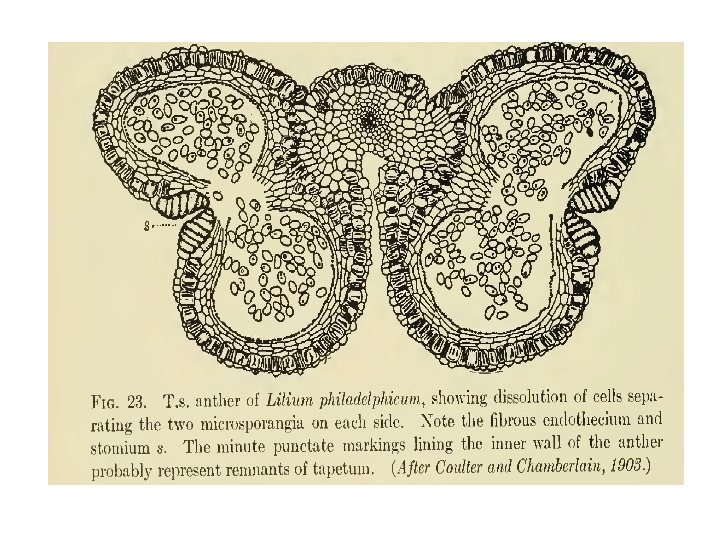
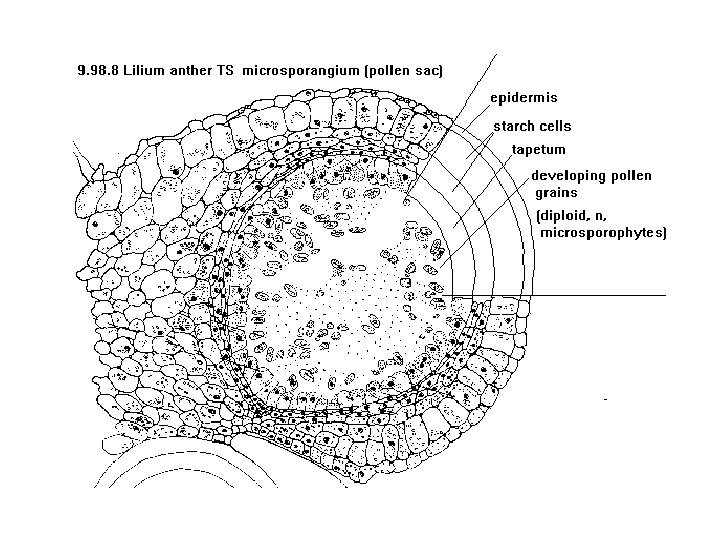
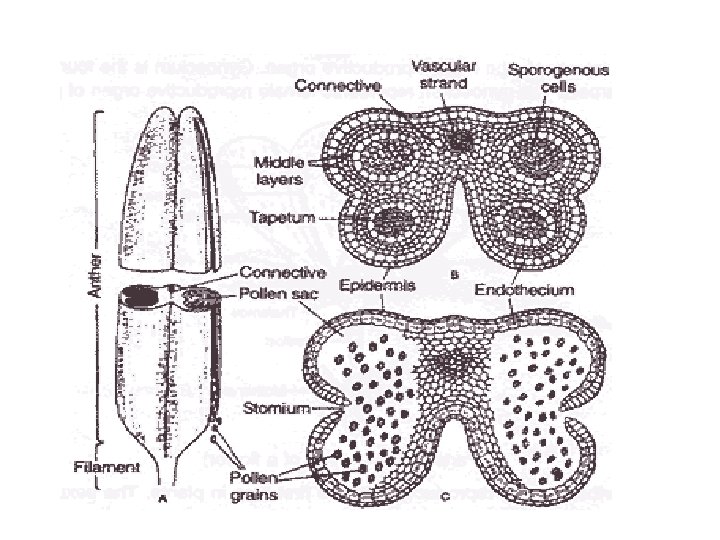
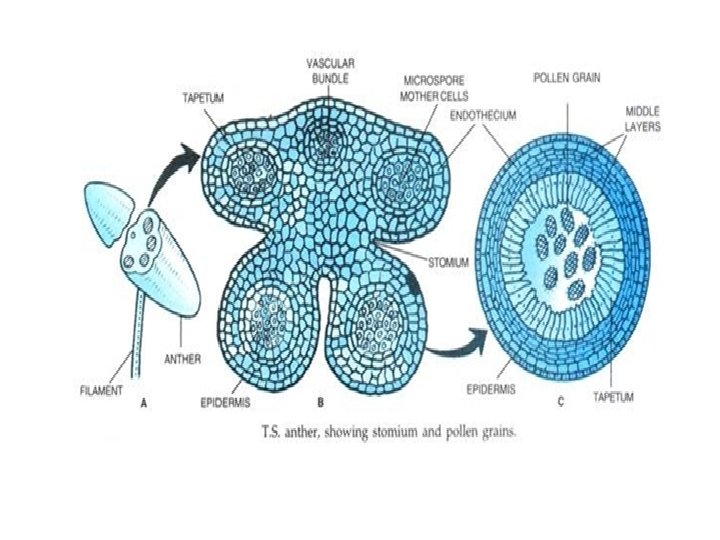
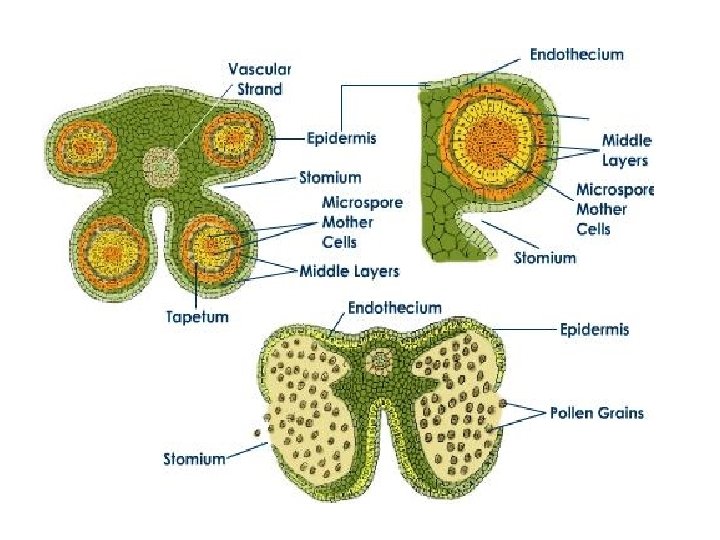
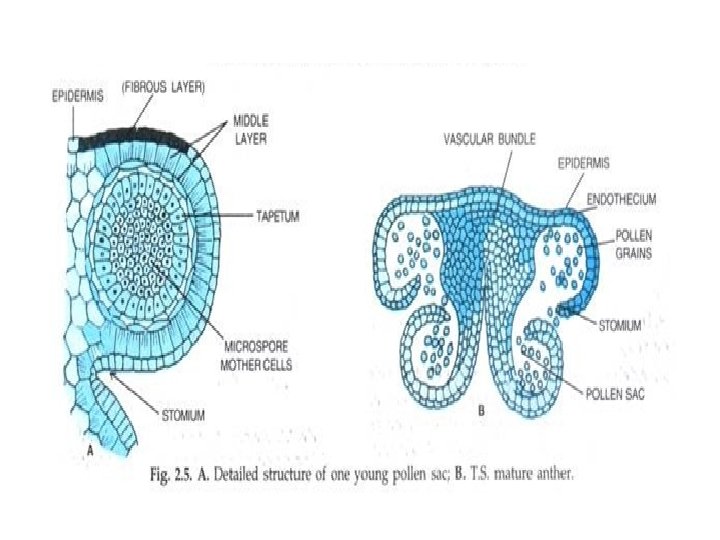
Microsporogenesis and development of the male gametophyte • Pollen is produced within the anthers (microsporangia or pollen sacs) of the flower. • During its development from an undifferentiated mound of cells (anther primordium) the anther forms two general groups of cells. • The reproductive or sporogenous cells give rise to the microspores and are formed from cells located centrally within the developing anther. •

• The non reproductive cells form discrete anther tissues layers and include the epidermal, cortical and tapetal cell layers surrounding the sporogenous cells. • The tapetum which is the innermost layer of the pollen sac plays a dominant role particularly during the microspore stage. • For example, many male sterile mutations affect tapetal cell functions and development is often arrested during the microspore stage.

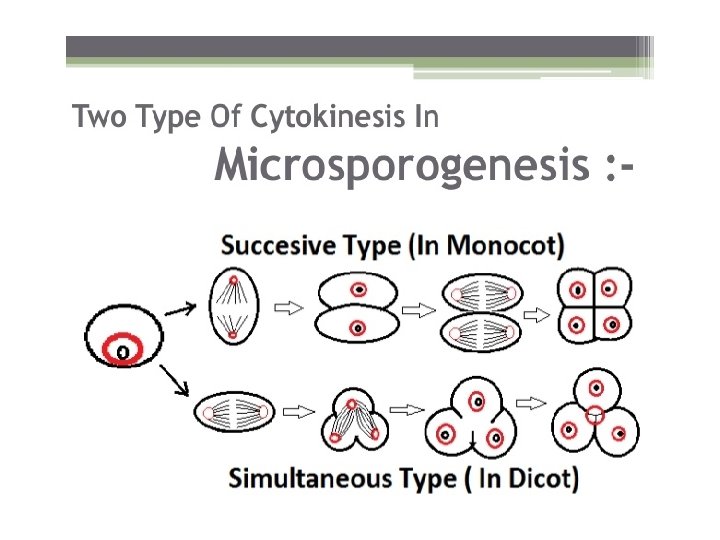
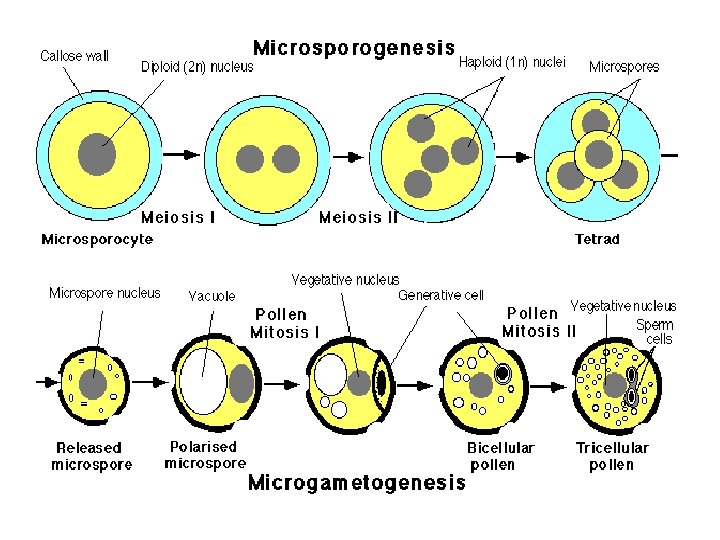
• Microsporogenesis comprises the events which lead to the formation of the haploid unicellular microspores. • During microsporogenesis the diploid sporogenous cells differentiate as microsporocytes (pollen mother cells or meiocytes) which divide by meiosis to form four haploid microspores. • Each diploid meiocyte gives rise to a tetrad of four haploid microspores and microsporogenesis is complete with the formation of distinct single celled haploid microspores.

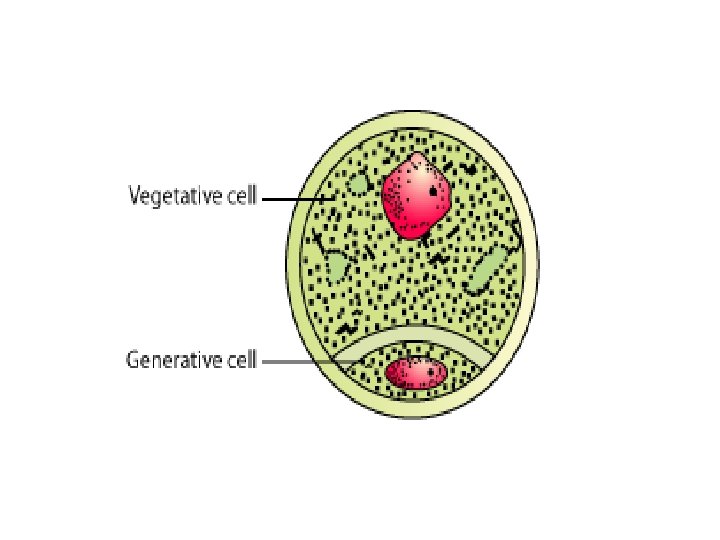
• The asymmetric cell division of the microspore at pollen mitosis I plays a critical role in the determination and subsequent fate of the two unequal daughter cells, the vegetative and generative cells. • As a result of this highly asymmetric division the larger vegetative cell does not divide further, accumulates a dense cytoplasm rich in protein, lipid and carbohydrates and possesses a large diffusely staining nucleus. • Storage compounds synthesised during vegetative cell maturation are utilised for intense metabolic activity during pollen tube growth by tip extension.

• In contrast, the smaller and metabolically repressed generative cell does not exit the cell cycle and continues through one more round of mitosis to produce two sperms cells with highly condensed chromatin. • This asymmetric division may be described as determinative in that daughter cells are immediately different (in size, gene expression and chromatin structure) after division and their fate is determined as a result of division asymmetry.
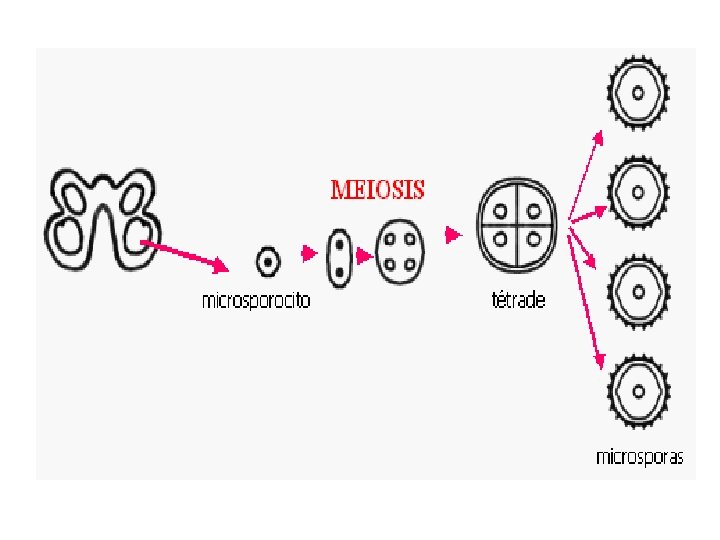

• Microgametogenesis comprises events which lead to the progressive development of the unicellular microspores into mature microgametophytes containing the gametes. • This phase begins with the expansion of the microspore which is commonly associated with the formation of a single large vacuole. • Vacuolation is accompanied by the displacement of the microspore nucleus to an eccentric position against the microspore wall. • In this position the nucleus undergoes first pollen mitosis (pollen mitosis I) which results in the formation of two unequal cells, a large vegetative cell and a small generative cell each containing a haploid nucleus.

• The generative cell subsequently detaches from the pollen grain wall and is engulfed by the vegetative cell forming a unique 'cell within a cell' structure. • The engulfed generative cell divides once more by mitosis (pollen mitosis II) to form the two sperm cells completely enclosed within the vegetative cell cytoplasm either before pollen is shed (tricellular pollen) or within the pollen tube (bicellular pollen).
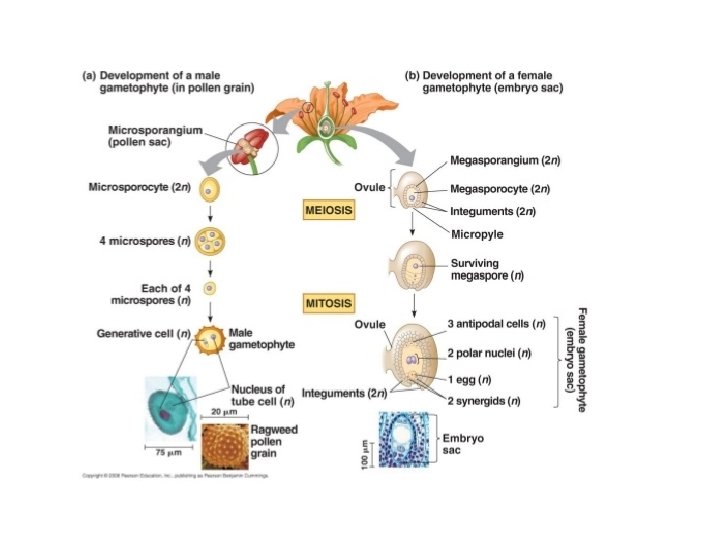

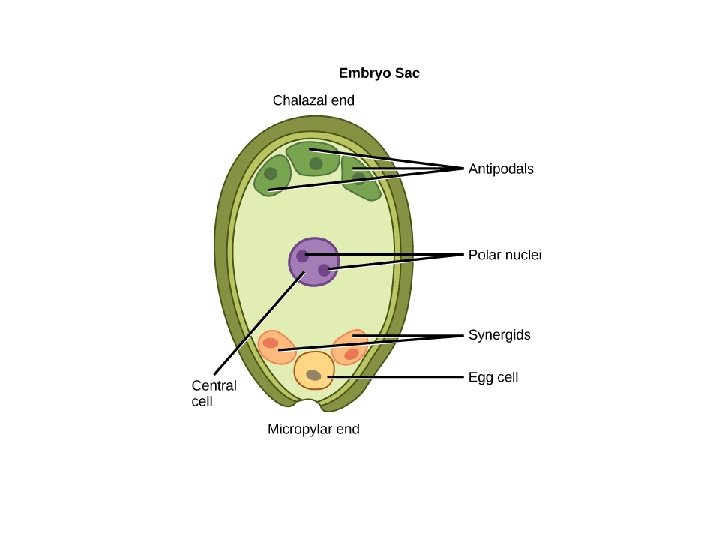
q Structure and development of Ovule: • Ovule means small egg. • In seed plants, the ovule is the structure that gives rise to and contains the female reproductive cells. • It consists of three parts: The integument(s) forming its outer layer(s), the nucellus and female gametophyte in its center. • The female gametophyte—specifically termed a "megagametophyte"—is also called the embryo sac in angiosperm. • The megagametophyte produces an egg cell (or several egg cells in some groups) for the purpose of fertilization. • After fertilization, the ovule develops into a seed

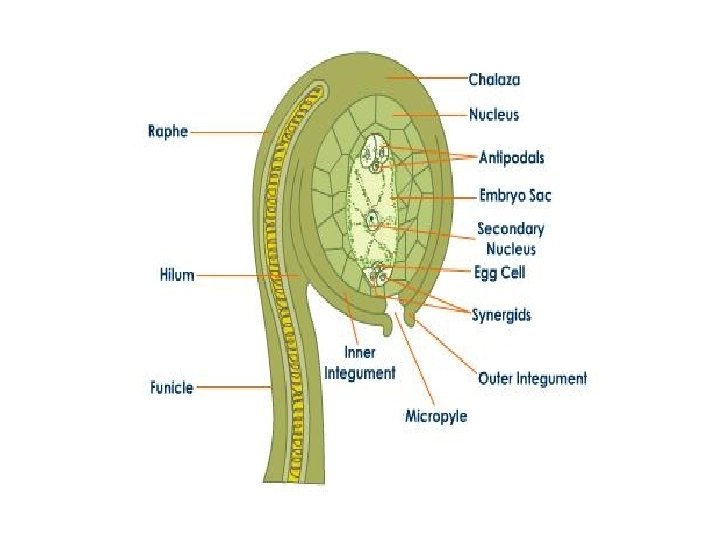

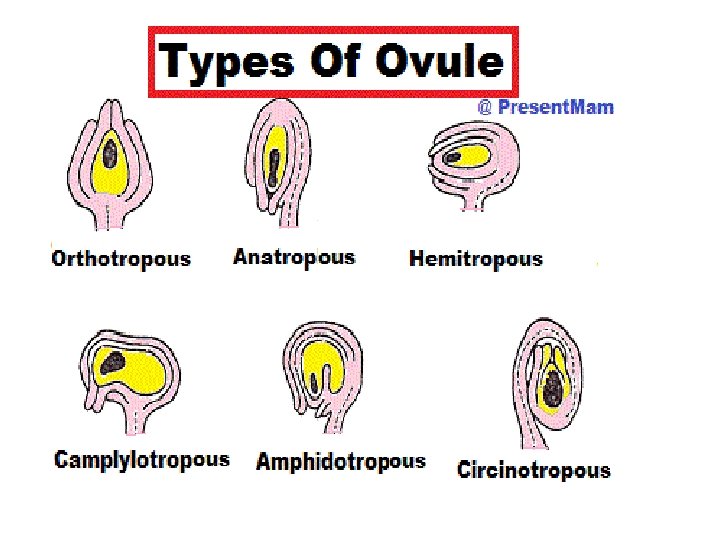
q Parts of Ovule: Ø Integuments: • An integument is a protective cell layer surrounding the ovule. • Gymnosperms typically have one integument (unitegmic) while angiosperms typically have two (bitegmic) Ø Micropyle: • The integuments do not enclose the nucellus completely but retain an opening at the apex referred to as the micropyle

Ø Chalaza: • Located opposite from the micropyle is the chalaza where the nucellus is joined to the integuments. • Nutrients from the plant travel through the phloem of the vascular system to the funiculus and outer integument and from there apoplast and symplast through the chalaza to the nucellus inside the ovule. Ø Nucellus: • The nucellus is a part of the inner structure of the ovule, forming a layer of diploid cells immediately inside the integuments. • It is structurally and functionally equivalent to the megasporangium.

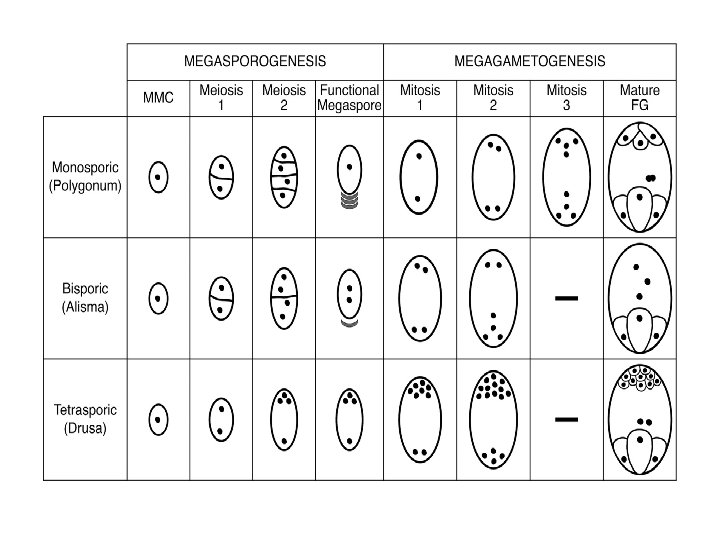
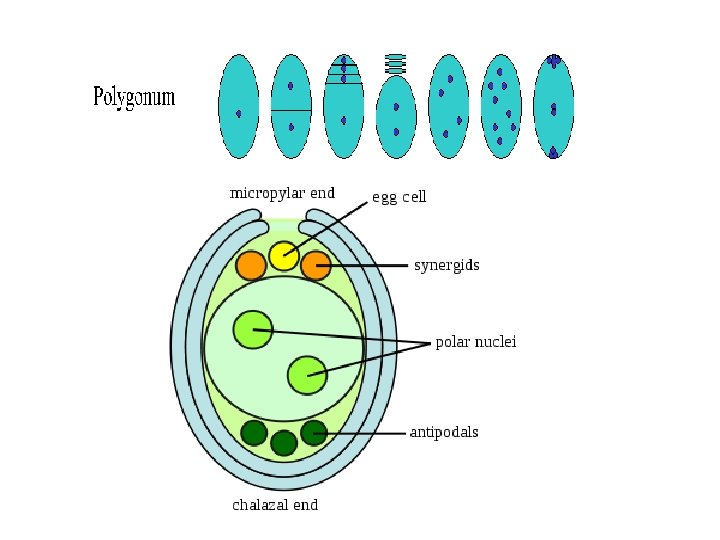
Ø Megaspore: • A common pattern of embryo sac development (the Polygonum type maturation pattern) includes a single functional megaspore followed by three rounds of mitosis. • In some cases, however, two megaspores survive (for example, in Allium and Endymion). • In many cases, all four megaspores survive. For example in Fritillaria type embryo sac development (illustrated by Lilium in the figure) three of the megaspores fuse to form a triploid nucleus. The subsequent arrangement of cells is similar to the Polygonum pattern, but the ploidy of the nuclei is different. Ø Perisperm: • After fertilization, the nucellus may develop into the perisperm that feeds the embryo.
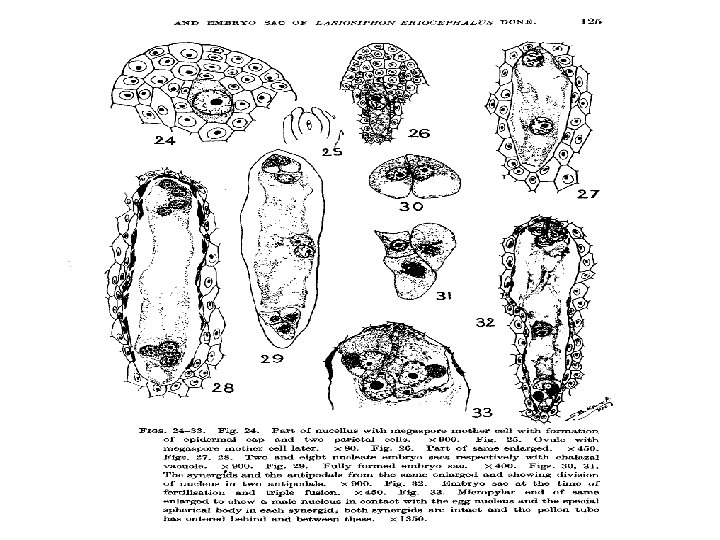

q Development of female gametophyte:

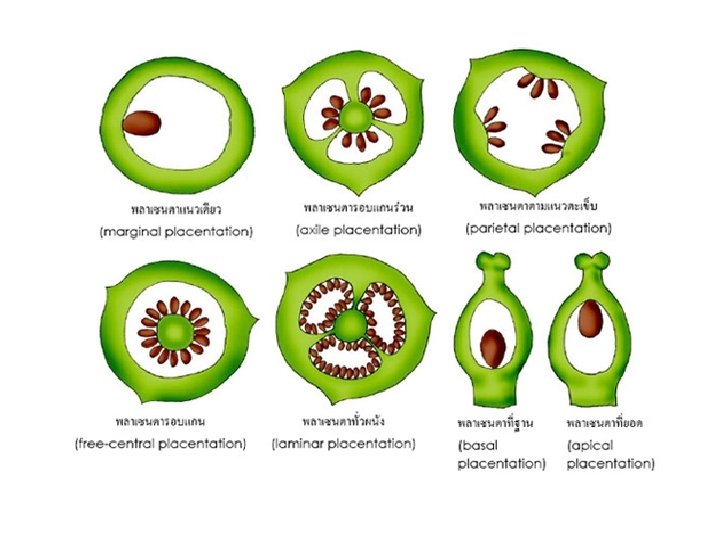
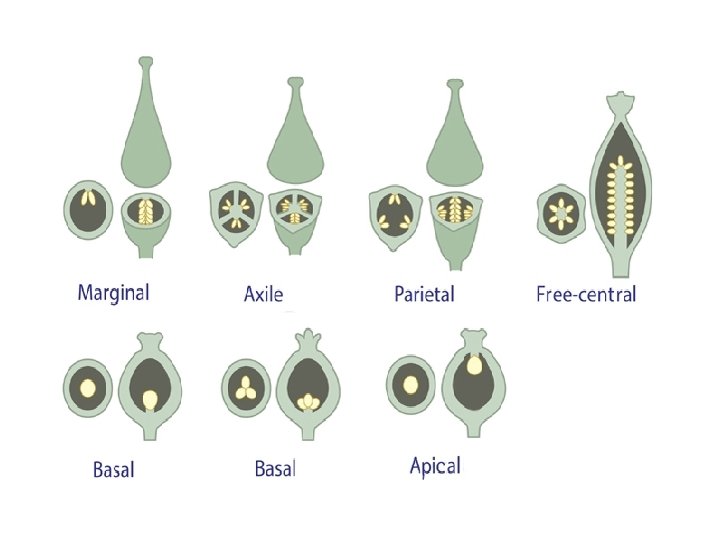
q. Placentation: The part of the ovary where the funiculus attaches is referred to as the placenta. • Basal placentation: The placenta is at the base (bottom) of the ovary. Simple or compound carpel. • Apical placentation: The placenta is at the apex (top) of the ovary. Simple or compound carpel. • Parietal placentation: The placentas are in the ovary wall within a non sectioned ovary. Compound carpel.

• Axile placentation: The ovary is sectioned by radial spokes with placentas in separate locules. • Free or central placentation: The placentas are in a central column within a non sectioned ovary. Compound carpel. • Marginal placentation: There is only one elongated placenta on one side of the ovary, as ovules are attached at the fusion line of the carpel's margins . This is conspicuous in legumes. Simple carpel.

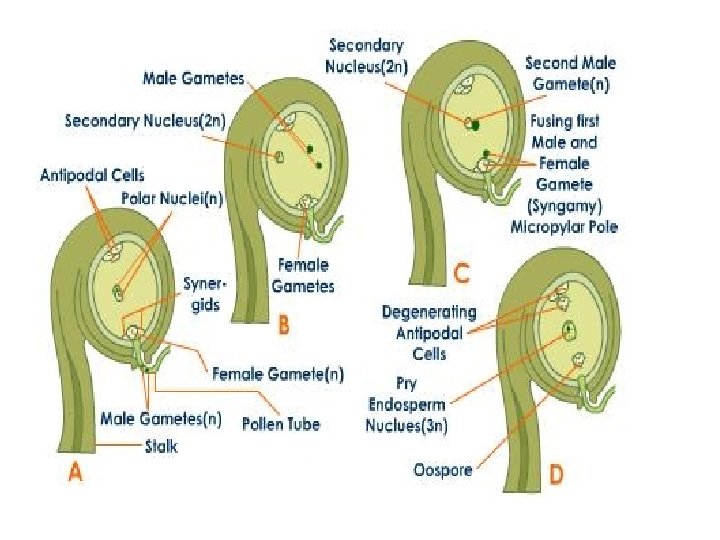
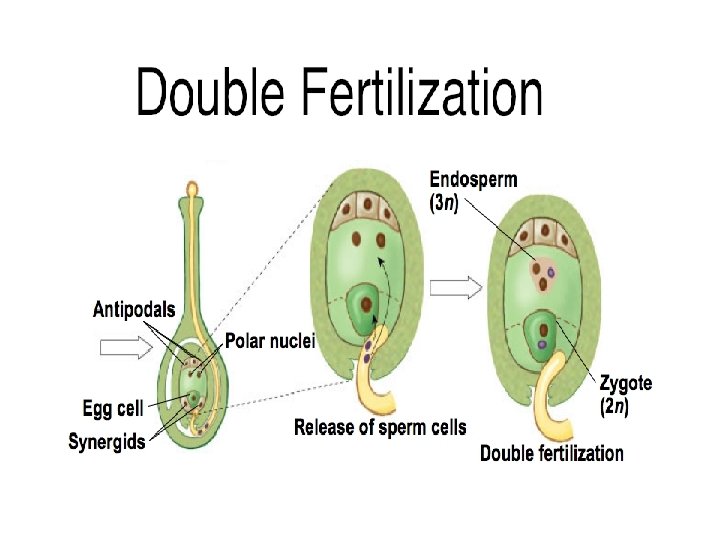
q. Fertilization: • The fusion of male and female gametes is called fertilization. • Fertilization in Angiosperms involves two nuclear fusions, one between one of the male gametes and the egg and the other between the second male gamete and the secondary nucleus of the polar nuclei. • The first of these fusions called generative fertilization leads to the formation of a zygote

• The second known as vegetative fertili zation leads to the formation of a triploid primary endosperm nucleus. • The zygote goes on to give rise to the embryo and the primary endosperm nucleus to a triploid nutritive tissue, called endosperm: hence the term doubles fertilization. • It is found only in angiosperms where it was first discovered by Nawaschin in 1898 in Fritillaria and Lilium.

Ovule Nucellus Pollen tube Male nucleus + 2 polar nuclei = endosperm nucleus (3 n) Male nucleus + egg cell = zygote (2 n) Pollen tube entering micropyle

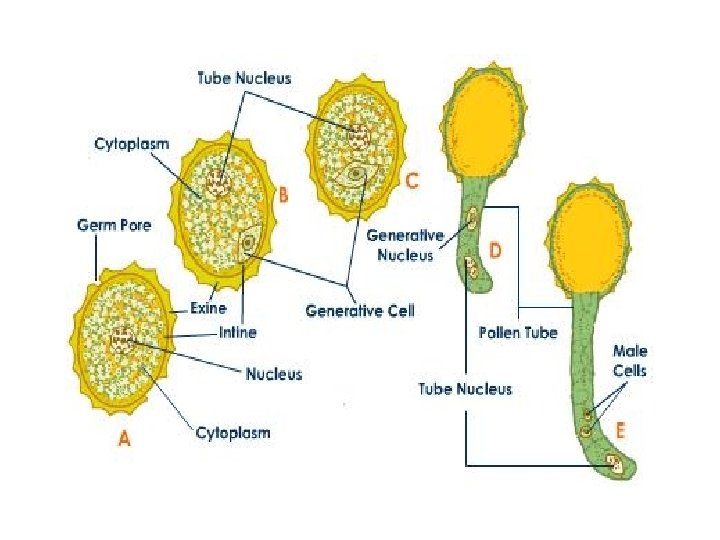
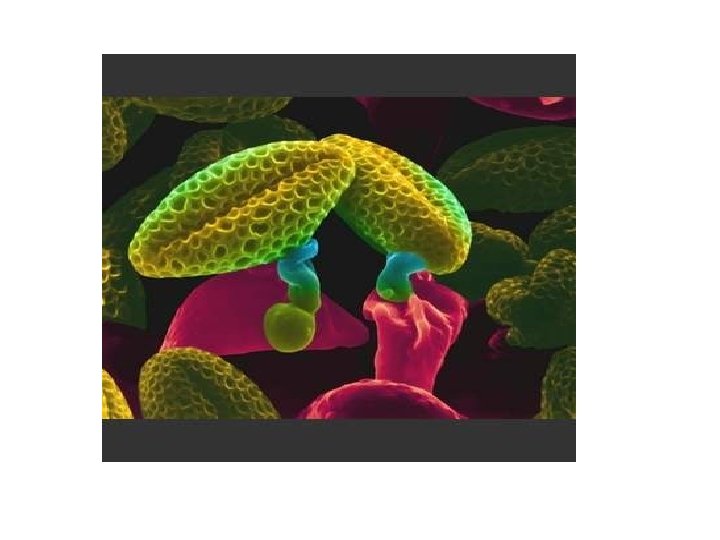
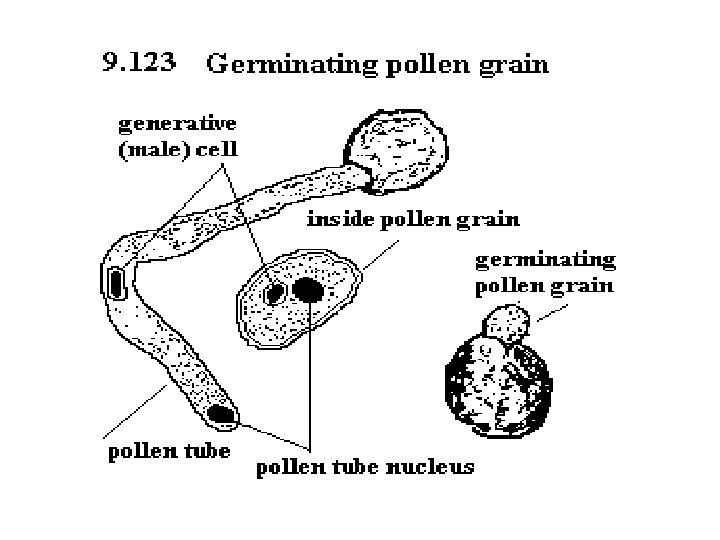
q Germination of Pollen: • A large number of pollen grains come to germinate over the papillose stigmatic surface. • On the stigma the pollen grain absorbs water and nutrients from the stigmatic secretion through its germs pores. • The tube or vegetative cell enlarges. It comes out of the pollen grain through one of the germ pores or germinal furrows to form a pollen tube. • It secretes pectinases and other hydrolytic enzymes to create a passage for it in the style if the latter is solid. The pollen tube absorbs nourishment from the cells of the style for its growth.

• The tube nucleus descends to the tip of the pollen tube. The generative cell (or its products) also passes into it. It soon divides into two male gametes if it is not already divided. • Each male gamete is lenticular to spherical in outline. It has a large nucleus which is surrounded by a thin sheath of cytoplasm. The tube nucleus may degenerate completely. • The pollen tube contains dense cytoplasm only towards it tip which also contains two male gametes and a degenerated tube nucleus. • Behind the tip the cytoplasm becomes highly vacuolate. It has plugs of callose for separating the older parts.

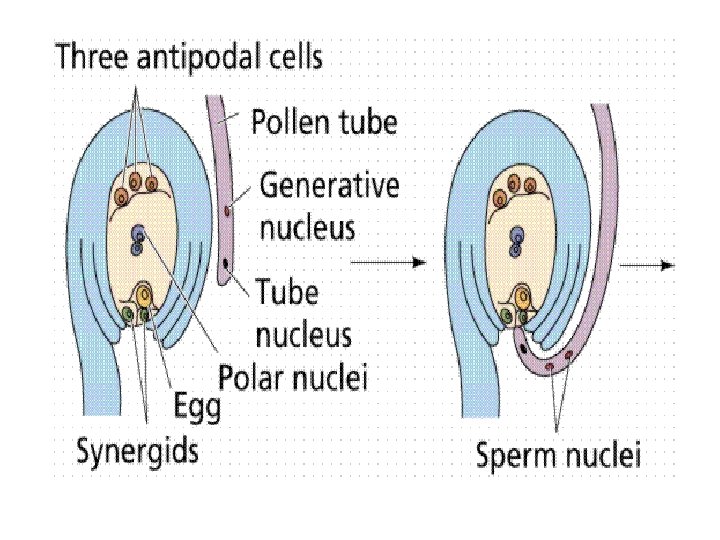
• In the ovary the growth of the pollen tube is directed by another tissue called obturator. • The pollen tube enters the ovule, either through its micropyle (porogamy e. g. , Lily), chalaza (chalazogamy e. g. , Casuarina, Juglans) or the sides after piercing through the integuments (mesogamy, e. g. Cucurbita, Populus). Porogamy is the most common. • After entering the ovule, the pollen tube is attracted towards the micropylar end of the embryo sac. The attractants are secreted by synergids or help cells. The pollen tube pierces one of the two synergids and bursts open into it.

• The synergid is simultaneously destroyed. Out of the two male gametes one fuses with egg or oosphere to perform generative fertilization. Generative fertilization is also called syngamy or true fertilization. • It gives rise to a diploid zygote or oospore. Soon after, the vacuole and plasmodesmal connections of the egg degenerate. • It now becomes ready to produce the embryo. • The nucleus of the second male gamete fuses with the two haploid polar nuclei or diploid secondary nucleus of the central cell to form a triploid primary endosperm nucleus.

• The central cell is now called primary endosperm cell. • This second fertilization is called vegetative fertilization since as a consequence of it a vegetative or nutritive tissue is formed to nourish the embryo. • Vegetative fertilization is also called triple fusion since three nuclei get fused, two polar nuclei and one male gamete.

q. Post Fertilization Changes: • The immediate consequence of double fertilization is the formation of zygote and the primary endosperm nucleus. • The zygote develops into the embryo (embryogeny) and primary endosperm nucleus into the endosperm (endosperm formation). • Besides these, there are many other macroscopic and microscopic changes that come about in the wake of fertilization. • The macroscopic changes, include the formation of a fruit and seed, general drying up of the flower and falling away of sepals, petals, stamens and styles. However, some of these organs might stay on and remain visible associated with the fruit.

q. The microscopic changes may be listed as follows: • 1. Zygote —> Embryo • 2. Primary endosperm nucleus —> Endosperm • 3. Integuments —> Seed coats • 4. Disintegration of nucellus; sometimes a small portion of it persist as a thin, papery covering, called perisperm in the seed. • 5. Conversion of the funicle into the seed stalk. • 6. Micropyle —> Persistent.
: • Embryogeny is the sum total of changes that occur")
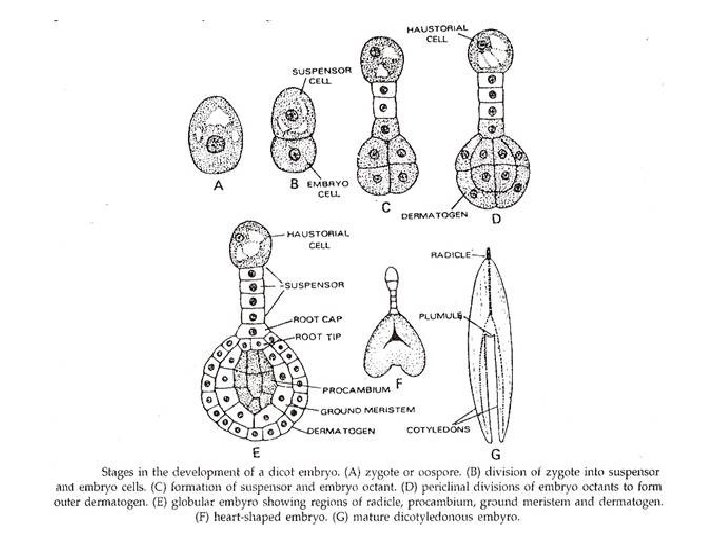
q. Embryogeny (Embryo Formation): • Embryogeny is the sum total of changes that occur during the development of a mature embryo from a zygote or oospore. • In a typical dicot, the zygote elongates and then divides by a transverse wall into two unequal cells. • The larger basal cell is called suspensor cell. The other towards the antipodal end is termed as terminal cell or embryo cell. • The suspensor cell divides transversely a few times to produce a filamentous suspensor of 6— 10 cells.
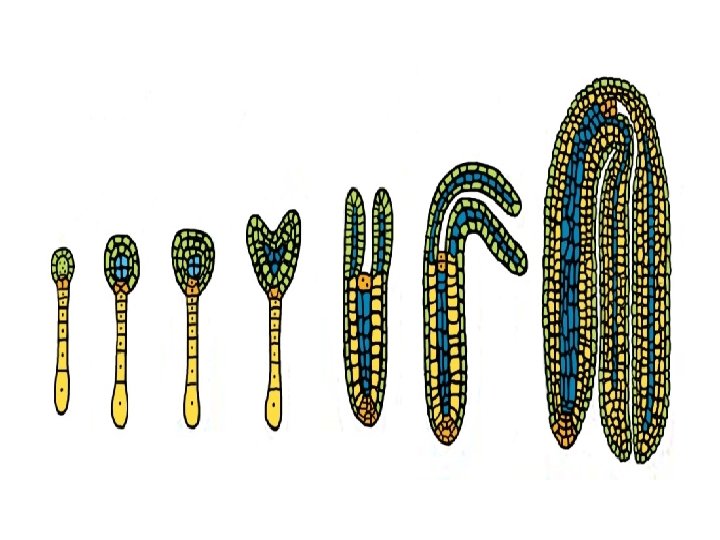
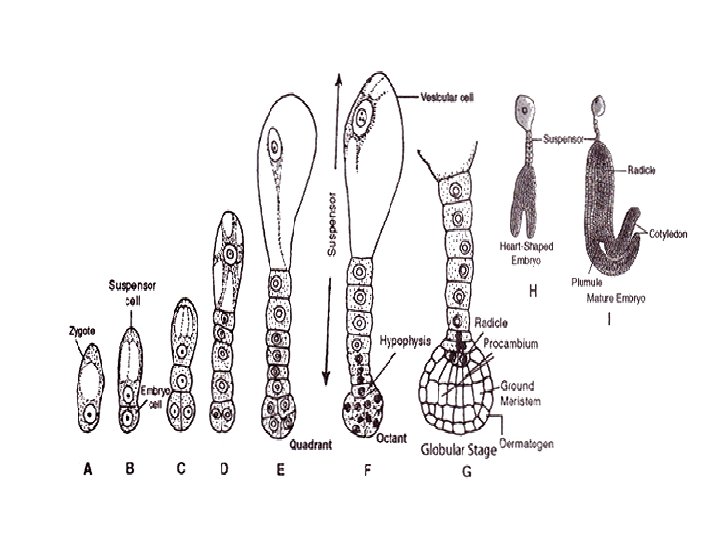

• The suspensor helps in pushing the embryo in the endosperm. • The first cell of the suspensor towards the micropylar end becomes swollen and functions as a haustorium. • The last cell of the suspensor at the end adjacent to the embryo is known as hypophysis. • Hypophysis later gives rise to the radicle.

• The embyro cell undergoes two vertical divisions and one transverse division to form eight cells arranged in two tiers—epibasal (terminal) and hypobasal (near the suspensor). • The epibasal cells eventually form the two cotyledons and the plumule. Only one cotyledon is produced in monocots (e. g. Wheat, Maize, Onion, Palm). The hypobasal cells produce the hypocotyl except its tip. • The eight embryonic cells or octants divide periclinally to produce an outer layer of protoderm or dermatogen.

• The inner cells differentiate further into procambium and ground meristem. • Proto derm forms epidermis, procambium gives rise to steal or vascular strand ground meristem pro duces cortex and pith.

q. Types of Endosperm in Plants: • Endosperm is the name of food laden tissue which is meant for nourishing the embryo in seed plants. • In gymnosperms it represents the female gametophyte hence haploid. • In angiosperms the endosperm is a special tissue which is formed as a result of vegetative fertilization, triple fusion or fusion of a male gamete with diploid secondary nucleus of the central cell. • The fusion product is primary endosperm cell having a triploid endosperm nucleus.
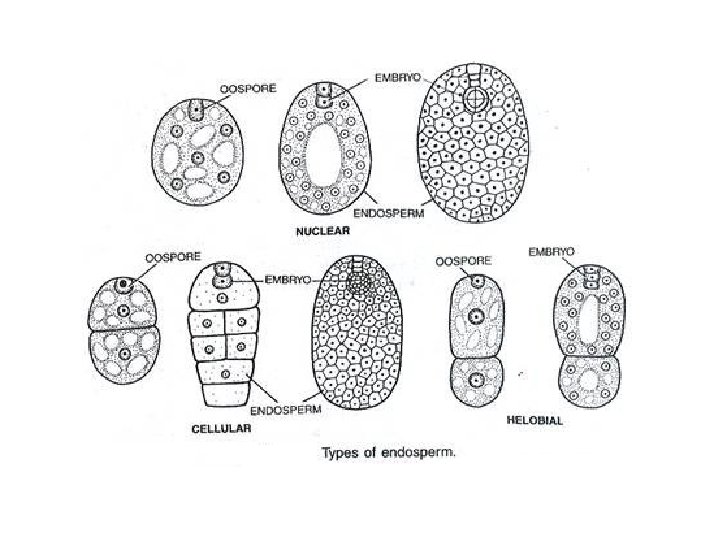

q Depending upon the mode of its formation, angiospermic endosperm is of three types: q Nuclear • The primary endosperm nucleus divides repeatedly without wall formation to produce a large number of free nuclei. • Meanwhile central vacuole appears in the central cell and pushes the cytoplasm containing the nuclei to the periphery. • The cytoplasm thickens so that the vacuole decreases in size. • It ultimately disappears with the exception of a few cases.

• The multinucleate cytoplasm undergoes cleavage and gives rise to a multicellular tissue, e. g. , Maize, Wheat, Rice and Sunflower. • In several cases, the cell wall formation remains incomplete. • For example, Coconut has multicellular endosperm (called coconut meal) in the outer part and free nuclear as well as vacuolated endosperm (called coconut milk) in the centre.

• In some cases wall formation occurs around the embryo only while in some it is restricted to the upper half. • Nuclear endosperm is the most common type of endosperm. • It is named so because it contains free nuclei in the beginning. q Cellular: • Every division of the primary endosperm nucleus is followed by cytokinesis. • Therefore, endosperm becomes cellular from the very beginning, e. g. , Bal sam, Datura, Petunia.

q Helobial: • It occurs in order helobiales of monocots. • The endosperm is of intermediate type between cellular and nuclear types. • The first division of primary endosperm nucleus is followed by transverse cytokinesis to form two unequal cells, larger micropylar and smaller chalazal. • Micropylar cell grows faster than the chalazal one. • Further development in both the cells occurs like that of nuclear endosperm, i. e. , multinucleate stage followed by wall formation, e. g. , Asphodelus. However, chalazal chamber often remains smaller and may degenerate.

q. Fate of Endosperm: • During its growth the endosperm crushes the nucellus. • It is in turn eaten by growing embryo. The endosperm may persist in the seed when the latter is called endospermic or albuminous (e. g. , Castor, Cereals, Coconut). • In others, the endosperm is completely absorbed by the growing embryo and the food reserve gets stored in the coty ledons. • Such seeds are called nonendospermic or exalbuminous, e. g. , Pea, Bean, Sunflower.
 and Passiflora, called ruminate endosperm.")
• Endosperm becomes convoluted in Areca (vem. Supari) and Passiflora, called ruminate endosperm. In Areca, Date (Pheonix) and Vegetable Ivory (Phytelephas microcarpa), the endosperm becomes very hard. The cellulosic hard endosperm of vegetable ivory is used to make buttons, umbrella handle heads and billiard balls.
- Slides: 83