Biological membranes Membranes Are Complex Structures Composed of

Biological membranes Membranes Are Complex Structures Composed of • Lipids, Proteins, & Carbohydrates. • Biological membranes consist of a double sheet (known as a bilayer) of lipid molecules, referred to as the phospholipid bilayer. Membranes also contains proteins and Carbohydrates.
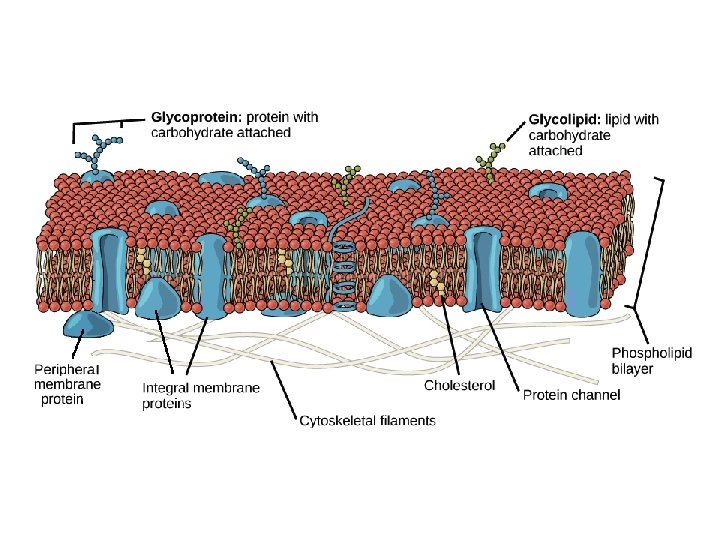

A-Lipids in membranes • Three types of lipid are found in biological membranes, namely phospholipids, glycolipids and sterols. • 1 -Phospholipids: include glycerophospholipids as phosphatidylcholine (PC), phosphatidylserine (PS), and phosphatidylethanolamine (PE), and sphingophospholipids as sphingomyelin. • 2 -Glycolipids: contain either glycerol or sphingosine have a sugar such as glucose in place of the phosphate head found in phospholipids. • 3 - Sterols: cholesterol in animal cell membranes

B-Carbohydrates • The sugars are attached to lipids and proteins can act as markers due to the structural diversity of sugar chains. For example, antigens composed of sugar chains on the surface of red blood cells determine an individual's blood group.

C-Proteins Membrane-spanning proteins are diverse in structure and function. They can be constructed of α-helices or from β-pleated sheets. Different Membranes Have Different Protein Compositions The number of different proteins in a membrane • varies from less than a dozen in the sarcoplasmic reticulum to over 100 in the plasma membrane,

• Most membrane proteins can be separated from one another using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), a technique that has revolutionized their study. Proteins are the major functional molecules of membranes and consist of enzymes, pumps and channels, structural components, antigens (eg, for histocompatibility), and receptors for various molecules. Because every membrane possesses a different complement of proteins, there is no such thing as a typical membrane structure

Amphipathic nature of membrane lipids • All membrane lipids are amphipathic—that is, they contain both a hydrophilic (water-loving) region and a hydrophobic (water-hating) region. Thus the most favourable environment for the hydrophilic head is an aqueous one, whereas the hydrophobic tail is more stable in a lipid environment. The amphipathic nature of membrane lipids means that they naturally form bilayers in which the hydrophilic heads point outward towards the aqueous environment and the hydrophobic tails point inward towards each other.


Subcellular organelles

1 -Mitochondria • Each mitochondrion is bounded by two membranes, which have very different functions and two separate mitochondrial compartments: the internal matrix and the intermembrane space. • a-The outer membrane contains many copies of a transport protein called porin, which forms large aqueous channels through the lipid bilayer.
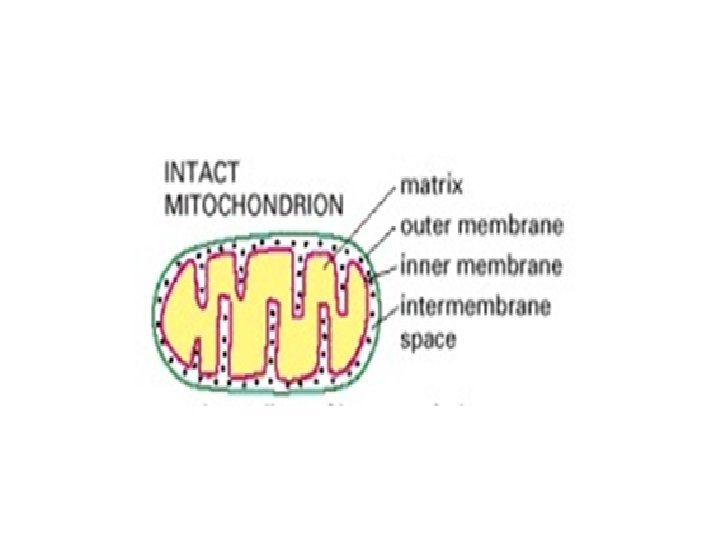

• b-The major working part of the mitochondrion is the matrix and the inner membrane. The inner membrane is usually highly convoluted, forming a series of infoldings, known as cristae, that project into the matrix. These convolutions greatly increase the area of the inner membrane, so that in a liver cell, for example, it constitutes about one-third of the total cell membrane. The number of cristae is three times greater in the mitochondrion of a cardiac muscle cell than in the mitochondrion of a liver cell, presumably because of the greater demand for ATP in heart cells.

• The matrix enzymes include those that metabolize pyruvate and fatty acids to produce acetyl Co. A and those that oxidize acetyl Co. A in the citric acid cycle. The principal end-products of this oxidation are CO 2, which is released from the cell as waste, and NADH, which is the main source of electrons for transport along the respiratory chain—the name given to the electrontransport chain in mitochondria. The enzymes of the respiratory chain are embedded in the inner mitochondrial membrane, and they are essential to the process of oxidative phosphorylation, which generates most of the animal cell's ATP.

2 -Endoplasmic Reticulum ER is an extensive membranous network in the • eukaryotic cells that is continuous with the outer nuclear membrane and composed of ribosomestudded (rough) and ribosome-free (smooth) regions. The ER is responsible for the manufacture of the membranes and performs many other biosynethic functions. Ribosomes on the rough ER, are used to synthesize proteins which are destined to either be inserted into the membrane of the cell or transported out of the cell.

Differences between ribosomes lying on • the ER and those lying in the cytosol are in the destination the protein that is synthesized by the ribosome. With protein synthesis via ER ribosomes, proteins are synthesized as they normally would through translation of m. RNA. After the protein is synthesized, called "posttranslation", it enters the lumen of the endoplasmic reticulum.
 of the")
Although the secondary structure (i. e. beta • sheets and alpha helixes) of the newlysynthesized peptide forms almost instantaneously as it is translated, its tertiary structure does not begin to form until it is in the lumen, where it will begin to fold into its specific 3 -D conformation with the aid of chaperone proteins

Inside the lumen of the rough • endoplasmic reticulum, N- and O- linked glycosidic bonds are formed between amino acids and sugars, thus this process is called "glycosylation" and the newly formed protein is called "glycoprotein"

The smooth endoplasmic reticulum, Its functions range from carbohydrate metabolism, lipid synthesis, and drug detoxification. Drug detoxification is usually accomplished by adding hydroxyl groups to drug molecules. This makes them more soluble in water and therefore easier to be flushed from the body. Another important function of the smooth endoplasmic reticulum is the storage of calcium ions. This is imperative for muscle contraction. When a muscle cell is stimulated by a nerve impulse, the calcium ions rush into the cytosol and trigger muscle contraction. Structurally the smooth ER is not continuous with the cell's nuclear envelope. The smooth ER is involved in phospholipid synthesis •

3 - Golgi • Extensive post-translational modification of membrane proteins undergo in Golgi apparatus. Many Golgi reactions involve the addition of sugar residues to membrane proteins and secreted proteins. The carbohydrates that the Golgi attaches to membrane proteins are often quite complex, and their synthesis requires multiple steps. Vesicles that bud off from the ER fuse with the closest Golgi membranes, called the cis-Golgi. •

Molecules then travel through the Golgi • apparatus via vesicle transport until they reach the end of the assembly line at the farthest sacs from the ER — called the trans-Golgi. At each workstation along the assembly line, Golgi enzymes catalyze distinct reactions. Later, as vesicles of membrane lipids and proteins bud off from the trans-Golgi,

4 -Lysosomes break down macromolecules into their constituent parts, which are then recycled. These membrane-bound organelles contain a variety of enzymes called hydrolases that can digest proteins, nucleic acids, lipids, and complex sugars. The lumen of a lysosome is more acidic than the cytoplasm. This environment activates the hydrolases and confines their destructive work to the lysosome. In plants and fungi, lysosomes are called acidic vacuoles. Lysosomes are formed by the fusion of vesicles that have budded off from the trans-Golgi.
- Slides: 21