Antibody Structure and Function Immunoglobulins are expressed as

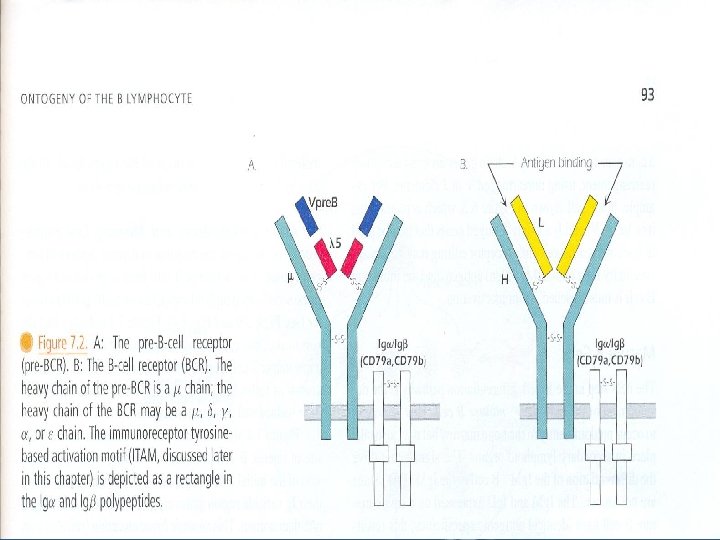
Antibody Structure and Function • Immunoglobulins are expressed as secreted and membrane-bound forms. • Membrane-bound antibody is present on the surface of B cells, it serves as antigen receptor. • Plasma cells produce secreted antibodies. • The membrane-bound form of antibody is associated with heterodimer called Ig /Ig to form the B cell receptor. • Ig /Ig heterodimer mediates the intracellular signaling mechanisms associated with B cell activation. • Antibodies exhibit specificity and biological activity.

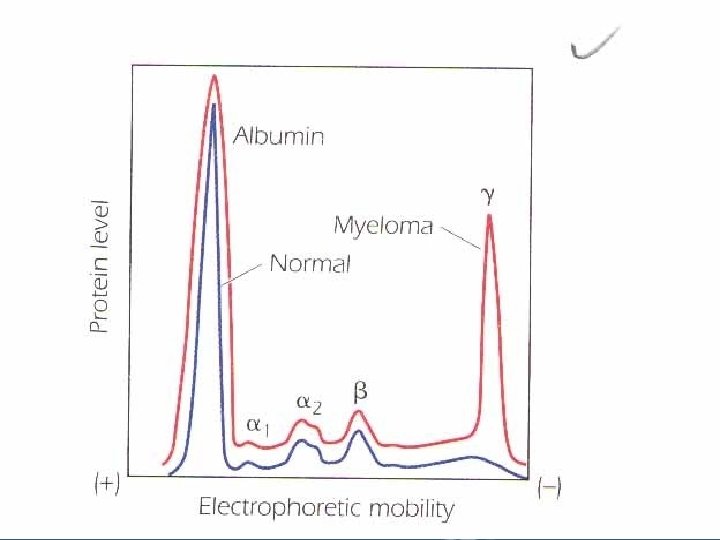
• The biological effects of antigen-antibody reactions include: – Neutralization of toxins. – Immobilization of microorganisms. – Neutralization of viral infectivity. – Agglutination of microorganisms. – Precipitation of soluble antigens. – Activation of serum complement to facilitate lysis of microorganisms. – Ability to cross placenta from mother to fetus. • The differences in biologic activities of antibodies are attributed to their isotypic "class" structure. • Antibody molecules are contained in the gamma globulin fraction when serum proteins are subjected to electrophoresis in slightly alkaline conditions "p. H 8. 2".

Structure of Light and Heavy Chains
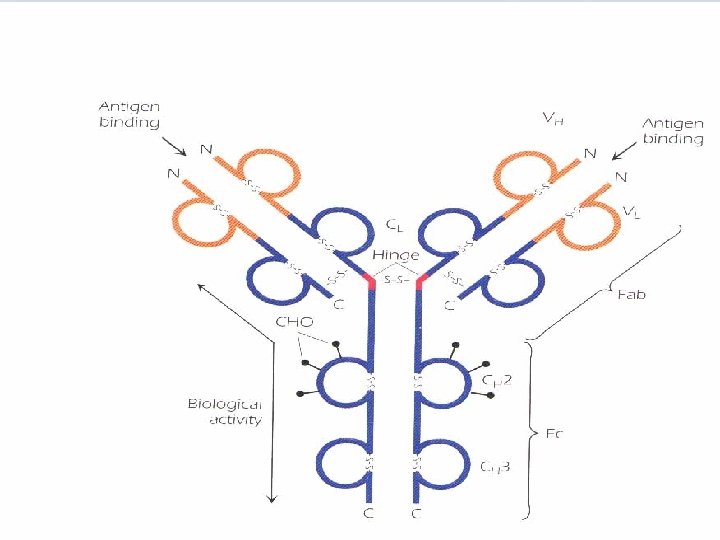

A. Papain split Ig molecule at the hinge region into 3 fragments of equal size "Fab + Fc" Fab fragments can bind antigen, but no precipitate is formed as they are univalent. FC is responsible for the biologic functions. B. Treatment with mercaptoethanol "breaks s-s bonds" four chains are obtained: • Two identical light chains "L". • Two indentical heavy chains "H". C. The antibody molecule is formed of 4 polypeptide chains, 2 L, 2 H which are linked by a number of disulfide bonds. D. Pepsin digestion results in formation of a divalent fragment F(ab`)2 consisting of two Fab regions joined by the disulfide bond, in addition several FC subfragments are obtained.

E. Other molecules belonging to immunoglobulin superfamily share this structural feature. • The light chains are always either both or both . • According to the structure of H chain there are five different classes "isotypes", H chains differ as antigens, carbohydrate content, and in size. H chains confer different biologic functions on each isotype. • in any one antibody molecule, both H chains are identical e. g. Ig. G class could have the structure 2 2 or 2 2 with two identical L chains and two identical H chains. • Some Ig have subclasses e. g. Ig. G subclasses are Ig. G 1, Ig. G 2, Ig. G 3, Ig. G 4. Ig. A 1 and Ig. A 2 Subclasses differ in the number and arrangement of interchain disulfide bonds and other structural features.

• L chains have two globular domains each, and H chains have 4 or 5 domains, separated by a short unfolded stretch. The first domain in L and H chains is highly variable, in terms of amino acid sequence, from one antibody to the next, it is designated VL or VH. The second and subsequent domains are on both H and L chains are constant and are designated CL and CH 1, CH 2, CH 3. In addition to their interchain disulfide bonding, the globular domains bind to each other in homologous pairs, by hydrophobic interactions as follows: VHLH, CH 1 CL, CH 2, CH 3.

The Hinge region is composed of a short segment of amino acids and is found between the CH 1 and CH 2 regions. This segment is made up of cysteine and proline residues. The cysteines are involved in formation of interchain disulfide bonds, and the proline residues prevent folding in a globular structure. The hinge region permits flexibility between the two Fab arms of the Y-shaped antibody molecule. It allows the two Fab arms to open and close to accommodate binding to two epitopes, separated by a fixed distance, as might be found on the surface of a bacterium.

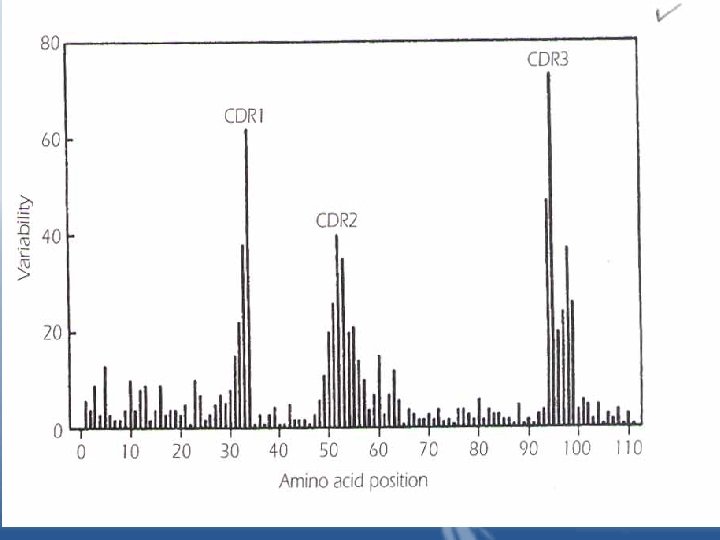
Variable region: • The greatest variability in sequence existed in the Nterminal 110 amino acids of both the L and H chains. • It occurs in 3 regions of the L and H chains, these regions are called hypervariable regions. • The hypervariable region participate in the binding with antigen and form the region complementary in structure to the antigen epitope. Hypervariable regions are termed "complementaritydetermining regions of the L and H chains: CDR 1, CDR 2, CDR 3". • Two dimensional model of the peptide chains are brought together in the folded form of the intact antibody molecule. Together, they constitute the combining site, which is complementary to the epitope.

• The variability in these CDRs provides the diversity in the shape of the combining site that is required for the function of antibodies of different specificities. • All forces involved in antigen-antibody interactions are weak, non-covalent interactions. • It is possible for two antibodies with different amino acid sequences to have specificity to the same epitope, in this case the binding affinities of the antibodies with the epitope will probably be different because of the difference in the number and types of binding forces.
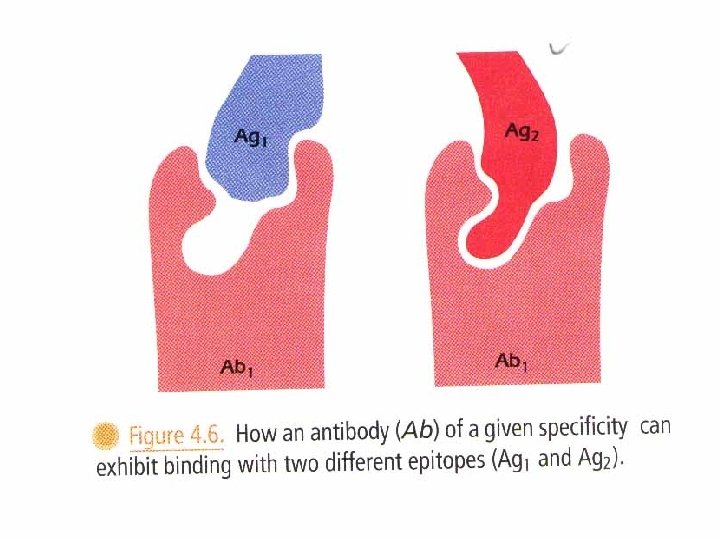

• Cross reacting compounds share antigenic determinants. • Some cross-reacting substance are unrelated to each other except that they have one or more epitopes in common. These substances are referred to as heterophile antigens. e. g. human blood group B antigen reacts with antibodies to certain strains of E. coli. The antigens of the microorganisms are referred to as heterophile antigens with respect to the blood group antigen.

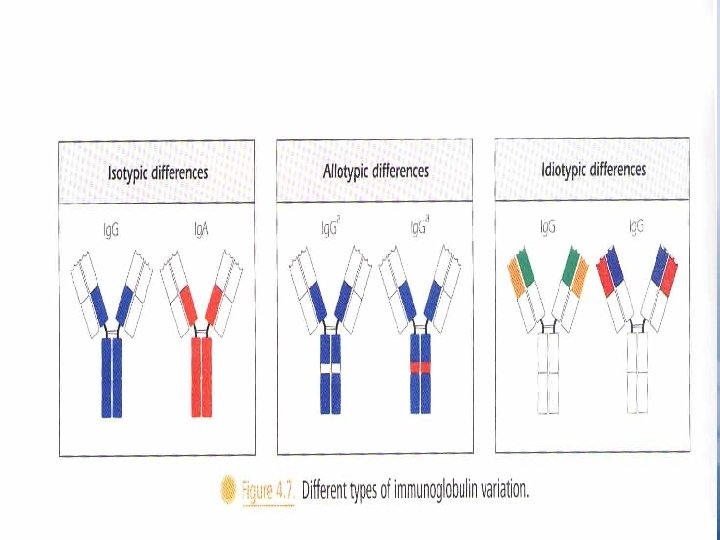
Immunoglobulin Variants Isotypes "fig. 4. 7" Allotypes • It is based on genetic differences among individuals, it depends on the existence of allelic forms "Allotypes" of the same protein, as a result of the presence of different forms of the same gene at a given locus. • Allotypic differences at known loci usually involve changes in only one or two amino acids in the constant region of a chain. "Fig 4. 7" • Allotypic bind the same antigen in similar way. • Allotypes markers e. g. Gm for chain of molecule Ig. G Km on the chain. AM on the chain • Allotypes are inherited as dominant Mendelian traits.

Idiotypes • Idiotypes represent differences in VL and VH region of the Ab used for inoculation. • They are immunogenic and capable of stimulating an immunologic response against itself in an animal of the same species. • In some case anti-idiotypic sera prevent binding of the Ab with its Ag. • Idiotypic determinants on Ig molecules may have a role in the control and modulation of immune response.

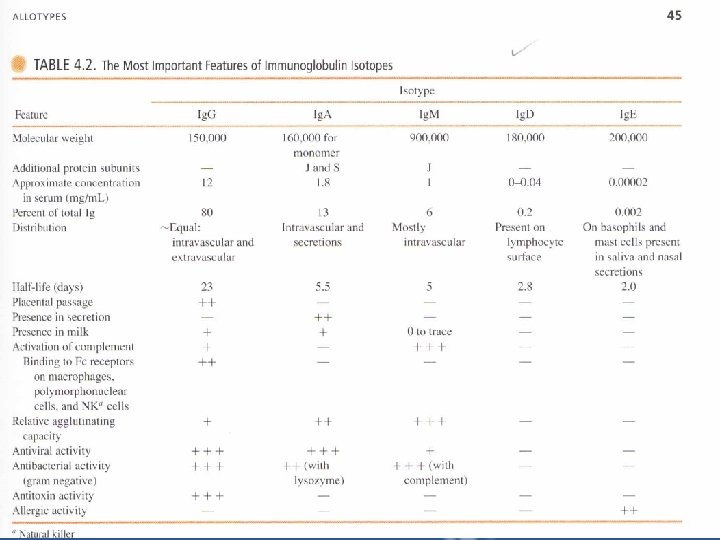
Ig. G – The predominant Ig in blood, lymph, CSF and pertinoeal fluid. It is distributed equally between the intravascular and extravascular spaces. • It consists of two heavy chains and two light chains "either or ". – There are four subclasses "Ig. G 1, Ig. G 2, Ig. G 3, Ig. G 4". Ig. G 1 is the most abundant. Ig. G subclasses differ in their chemical and biologic properties. – Figure " page 12 " explains why the half life of Ig. G is longer than other classes. – Ig. G molecules can cause the agglutination or clumping of particulate "insoluble" Ags such as microorganisms. – The reaction of Ig. G with soluble, multivalent Ags can generate precipitates. Insoluble Ag-Ab complexes are easily phagocytized and destroyed by phagocytic cells.

– Ig. G crosses the placenta The Ig. G isotype (except for Ig. G 2) is the only class of Ig that can pass through placenta, enabling the mother to transfer her immunity to the fetus. • Placental transfer is facilitated by expression of an Ig. G protection receptor "Fc. Rn" expressed on placental cells. Fc. Rn are identical to the Ig. G protection receptor "Fc. Rp" found in the cellular endosomes. • At the 3 rd or 4 th month of the pregnancy, there is rapid increase in the concentration of Ig. G(of maternal origin). • During 5 th month of pregnancy, the fetus begins to make Ig. M and trace amount of Ig. A.

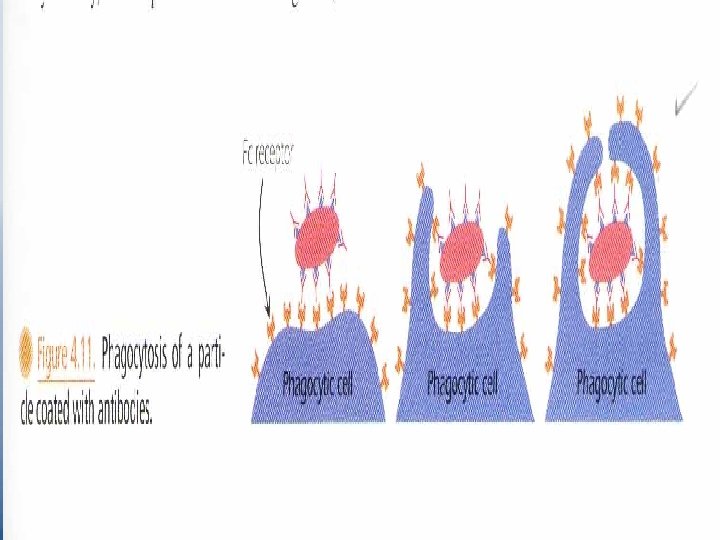
• Three or four months after birth, the infant begins to synthesize its own Ig. G which coincides with the drop of inherited maternal Ig. G. • Absorption of Maternal Ig. G contained in colostrums of nursing mothers is achieved by its binding Fc. Rn protection receptor expressed in the intestinal tissue of neonates. § Oposonization is a property of Ig. G; it facilitates phagocytosis. • Phagocytes express membrane receptors for the Fc portion of the Ig. G molecule. These cells adhere to the Ab-coated bacteria by virtue of their Fc receptors.

– Antibody-dependent, cell-Mediated cytotoxicity "ADCC". The Fab portion of Ig. G binds with the target cell "microorganism or tumor cell" and the Fc portion binds receptors on NK cells. Ig. G thus focuses the killer cells on their target, and the killer cells destroy the target by releasing certain substances. – activation of complement by Ig. G complement activation leads to the generation of components that cause opsonization, phagocytosis and direct lysis of the invading organism. – Neutralization of Toxin Ig. G blocks the active site of various toxins "tetanus, botulinus …", passive immunization is used. – Immobiliztion of Bacteria e. g. TPI. – Neutralization of viruses Ig. G is an efficient virus neutralizing Ab. Inhibition of viral attachment to the target cell, or inhibition of viral penetration.


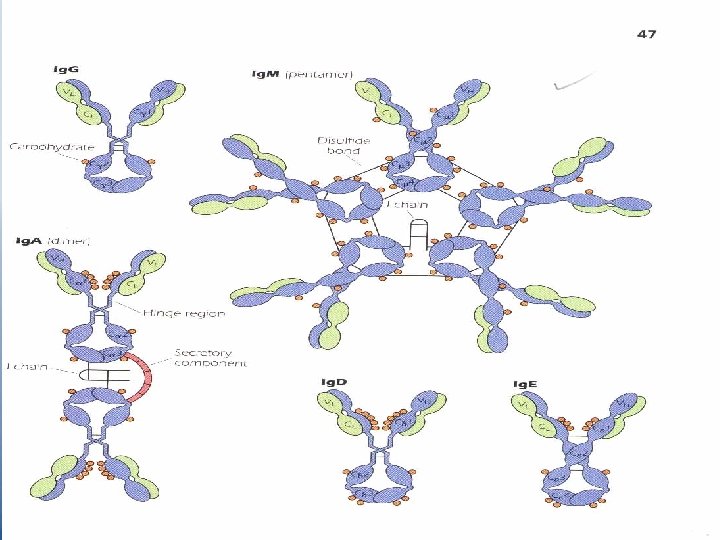
Ig. M – The first Ig produced following immunization elevated levels indicate recent infection or recent exposure to Ag. – Pentameric molecule, the units are joined together by additional s-s bonds between their Fc portions and by a polypeptide chain "J chain" which is synthesized in B or plasma cells. – Ig. M is found in human serum and intravaseular spaces. It is also found on the surface of mature B cells together with Ig. D, where it serves as an Ag-specific BCR. – Ig. M is an excellent complement-fixing. – Ig. M molecules are efficient agglutinating Abs. – Isohemagglutinins are naturally occurring Ig. M Abs against RBC Ags of ABO blood groups.

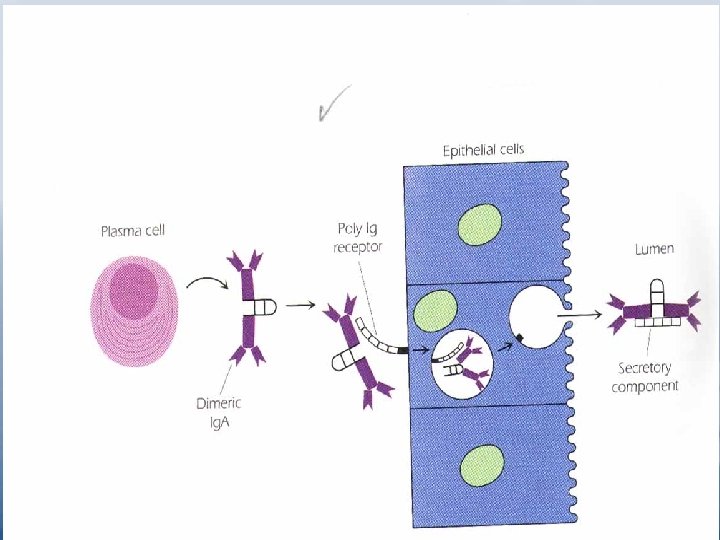
Ig. M isohemagglutinins do not pass through the placenta, so incompatibility of the ABO between mother and fetus poses no danger to the fetus. Some bacteria in the GIT and RT bear determinants similar to the oligosaccharides of the ABO blood groups. Ig. A • The major Ab in External secretions such as saliva, mucus, sweat, gastric fluid and tears. • The major Ab found in the colostrums of milk in nursing mother, it protects intestinal tract-of neonates against pathogens during the first few weeks after birth. • Within mucous secretions, Ig. A exists as a dimmer joined by J chain.

• When dimeric molecules are released from plasma cells, they bind to the poly-Ig receptor expressed on the basal membrance of adjacent epithelial cells. This receptor transports the molecule through the epithelial cells and releases them into extracellular fluids (e. g. in the gut or bronchi). Release is facilitated by enzymatic cleavage of the poly-Ig receptor, leaving a large fragment "the secretory component" of the receptor still attached to the Fc piece of the dimeric Ig. A molecule (Figure 4. 13 p. 52). The secretory component may help protect the dimeric Ig. A from proteolytic cleavage. Secretory Ig. A protects respirtory and GIT from infection. It may inhibit attachment of invading organisms to the epithelial surface. • Ig. A has bactericidal activity against gram-negative organisms in the presence of lysozyme. • Ig. A can act as antiviral and is an efficient agglutinating Ab.

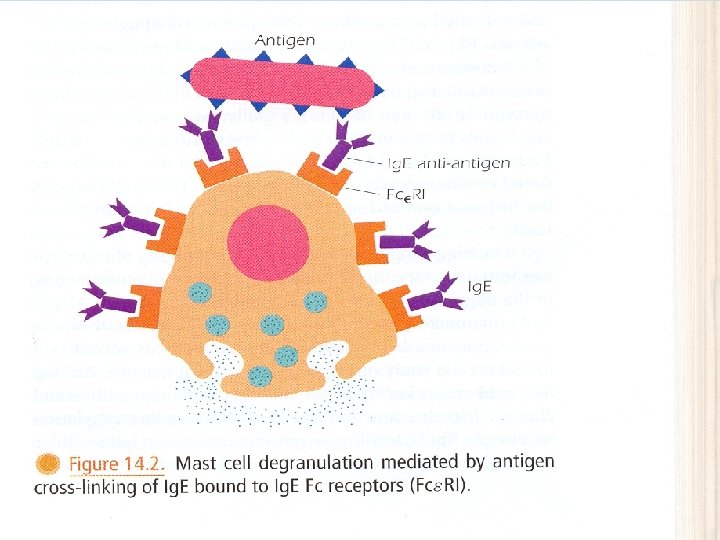
Ig. D • Ig. D is co-expressed with Ig. M on the surface of mature B cells. As Ig. M, it functions as an Ag-specific BCR. • Ig. D appears to correlate with the elimination of B cells with the capacity to generate self-reactive Abs. Ig. E • termed reaginic Ab. • Fc of Ig. E binds efficiently to Fc receptors on mast cells and basophils. • When Ag reappears, it combines with Fab of Ig. E attached to these cells, causing it to be cross linked, then the cell becomes activated and releases histamine, heparin, leukotrienes and triggers the immediate hypersensitivity reaction. • Ig. E has a role in protection against certain parasites.
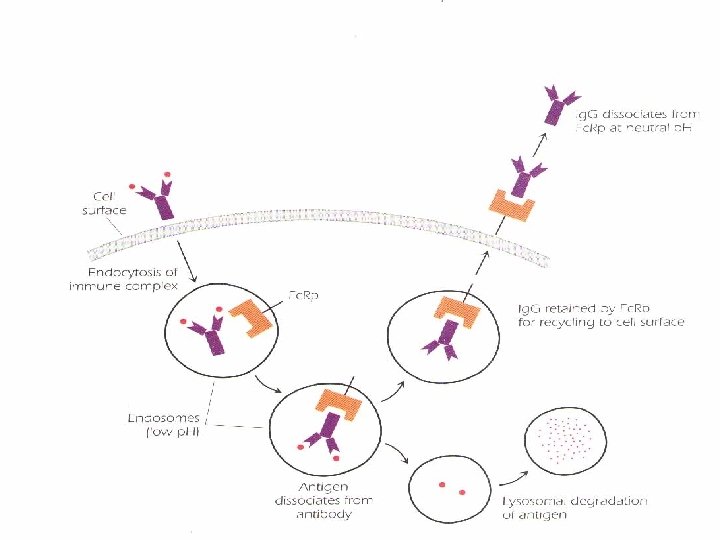
. Circulating monomeric Ig. G plus antigen{immune")
Recycling of Ig. G utilizing protector receptor(Fc. Rp). Circulating monomeric Ig. G plus antigen{immune complex) enters an antigen-presenting cell through the process of endocytosis. Whithin the endosome, the complex binds Fc. Rp; Ig. G and Ag dissociate, allowing the Ig. G to be directed to the cell surface for recycling. Antigen undergoes lysosomal degradation (Ag processing), and its proteolytic fragments are ultimately expressed on the cell surface in the context of MHC class II molecules

Kinetics of the Ab response after immunization

• Primary response. • The 1 ry Ab response is divided into several phases: • 1 - Latent or Lag period • After initial exposure to an Ag, a period of 1 -2 weeks follows before Ab is detectable in serum. It is the time taken for T and B cells to make contact with the Ag, to proliferate and differentiate • 2 - Exponential phase • The concentration of Ab in the serum increases exponentially. • 3 -Steady state • Production and degradation are balanced.

• 4 -Declining phase • Immune response shuts down, Ab level in serum declines rapidly. • The first class of Ab detected in 1 ry response is Ig. M, if Ig. G production ensues, its appearance is accompanied by a rapid cessation of production of Ig. M. • Secondary Response • Following 1 ry response, a pool of long-lived memory cells are left which are capable of mounting a secondary response as well as any other future responses to the Ag. • Secondary or memory response( also called anamnestic response) becomes apparent when a response is triggered by a second injection of the same Ag. • In the 2 ry immune response the lag phase is shorter, more Ab is produced which continue for longer period( months to years).

• Ig. G Abs appear at higher concentration and Ig. M levels drop, Ig. A and Ig. E may also appear( this shift is known as class switching) but all classes have the same Ag specificity as the original Ig. M. • Affinity maturation occurs( increased binding capacity of Abs for Ag). • 2 ry immune response justifies repeated immunization with the same attenuated or inactivated pathogens or their products.

Ig superfamilty

They share structural features with Ig • Ig-fold domains. • Genes encoding them arose from a common primordial gene. • Ig-fold structures "loops" formed as a result of intrachain disulfide bonds and consisting of approximately 110 amino acids. • These domains may facilitate interactions between membrane proteins "e. g. CD 4 on TH cells and MHC II on APC".

B- Cell Hybridomas and Monoclonal Antibodies

• The immunizing agent which usually has many epitopes leads to production of antisera that contains a mixture of anti bodies with varying specificity for all epitopes. • The reason is that various population of antibody forming lymphocytes are expanded. • In 1975 Kohler and Milstien developed methods for generation of monoclonal antibodies they shared the noble prize for this development. • Monoclonal antibodies are homogeneous populations of antibody molecules, derived from a single antibody-producing cell , in which all antibodies are identical and of the same precise specificity for a given epitope.

Procedure: 1. Malignant plasma cells "immortal in cell culture" that do not produce Ig are used. The cells are engineered to be deficient in an enzyme hypoxanthine guanine phosphoribosyl transferase " HGPRT", they will not survive in culture unless the enzyme is added to the media. 2. These cells are fused "hybridized" with a source of freshly harvested B cells from a mouse recently immunized with Ag " e. g. spleen cells" to form B cell hybridomas. The fusion is accomplished by polyethylene glycol " PEG" 3. The cells are then cultured in media lacking HGPRT. – Antibody- producing B cells produce HGPRT will survive. – Hybridoma cells will survive. – Within days the non – fused HGPRT negative plasma cells soon die as do all nonfused B cells.

– Those hybrid cells synthesizing specific antibody are selected by testing for Ag reactivity "e. g. ELISA". – Those cells are cloned from single cells each clone synthesizing Abs of a single specificity. Uses of MABS: • MABS are used for diagnostic tests, in immune therapy of cancer in which various drugs or toxins are conjugated to MABS which in turn deliver these substances to the tumor cells against which the Abs are specifically directed. •
- Slides: 44