5 PHYSIOLOGY OF PERIPHERAL NERVOUS SYSTEM PNS MODULE

")
 Inward current depolarizes the membrane potential to")
 The combined effect of closing the Na+ channels and greater opening of the")





 b.")



 increases")
 reflex—knee jerk It is monosynaptic. a. Muscle is")

. It results in flexion on the ipsilateral side and")


- Slides: 20
5. PHYSIOLOGY OF PERIPHERAL NERVOUS SYSTEM PNS MODULE Prof. Sherif W. Mansour Physiology dpt. , Mutah school of medicine. 2020/2021
Action potential It is a property of excitable cells (i. e. , nerve, muscle) that consists of a rapid depolarization, or upstroke, followed by repolarization of the membrane potential. Action potentials have stereotypical size and shape, are propagating, and are all-ornone. - Threshold : is the membrane potential at which the action potential is inevitable. At threshold potential, net inward current becomes larger than net outward current. The resulting depolarization becomes self-sustaining and gives rise to the upstroke of the action potential. If net inward current is less than net outward current, no action potential will occur (i. e. , all-or-none response). Ionic basis of the nerve action potential a. Resting membrane potential - It is approximately − 70 m. V, inside negative. - is the result of the high resting conductance to K+ > Na+ (100 times). At rest, the Na+ channels are closed and Na+ conductance is low.
b. Upstroke of the action potential (1) Inward current depolarizes the membrane potential to threshold. (2) Depolarization causes rapid opening of the activation gates of the Na+ channels, and the Na+ conductance of the membrane promptly increases. (3) The Na+ conductance becomes higher than the K+ conductance, and the membrane potential is driven toward (but does not quite reach) the Na+ equilibrium potential of +65 m. V. Thus, the rapid depolarization during the upstroke is caused by an inward Na+ current. (4) The overshoot is the brief portion at the peak of the action potential when the membrane potential is positive. c. Repolarization of the action potential (1) Depolarization also closes the inactivation gates of the Na+ channels (but more slowly than it opens the activation gates). Closure of the inactivation gates results in closure of the Na+ channels, and the Na+ conductance returns toward zero. (2) Depolarization slowly opens K+ channels and increases K+ conductance to even higher levels than at rest
(3) The combined effect of closing the Na+ channels and greater opening of the K+ channels makes the K+ conductance higher than the Na+ conductance, and the membrane potential is repolarized. Thus, repolarization is caused by an outward K+ current. d. Undershoot (hyperpolarizing after potential) - The K+ conductance remains higher than at rest for some time after closure of the Na+ channels. During this period, the membrane potential is driven very close to the K+ equilibrium potential.
Refractory periods a. Absolute refractory period It is the period during which another action potential cannot be elicited, no matter how large the stimulus. -It coincides with almost the entire duration of the action potential. ■ Explanation: Recall that the inactivation gates of the Na+ channels are closed when the membrane potential is depolarized. They remain closed until repolarization occurs. No action potential can occur until the inactivation gates open. b. Relative refractory period It begins at the end of the absolute refractory period and continues until the membrane potential returns to the resting level. -An action potential can be elicited during this period only if a larger than usual inward current is provided.
■ Explanation: The K+ conductance is higher than at rest, and the membrane potential is closer to the K+ equilibrium potential and, therefore, farther from threshold; more inward current is required to bring the membrane to threshold. c. Accommodation It occurs when the cell membrane is held at a depolarized level such that the threshold potential is passed without firing an action potential. -It occurs because depolarization closes inactivation gates on the Na+ channels. -It is demonstrated in hyperkalemia, in which skeletal muscle membranes are depolarized by the high serum K+ concentration. Although the membrane potential is closer to threshold, action potentials do not occur because inactivation gates on Na+ channels are closed by depolarization, causing muscle weakness.
Propagation of action potentials ■ occurs by the spread of local currents to adjacent areas of membrane, which are then depolarized to threshold and generate action potentials. ■ Conduction velocity is increased by: a. ↑ fiber size. Increasing the diameter of a nerve fiber results in decreased internal resistance; thus, conduction velocity down the nerve is faster. b. Myelination. Myelin acts as an insulator around nerve axons and increases conduction velocity. Myelinated nerves exhibit salutatory (jumping) conduction because action potentials can be generated only at the nodes of Ranvier, where there are gaps in the myelin sheath.
RECEPTORS The environmental signals that can be detected include mechanical force, light, sound, chemicals, and temperature. - Types of sensory receptors: a. Mechanoreceptors Pacinian corpuscles , Joint receptors, Stretch receptors in muscle, Hair cells in auditory and vestibular systems, Baroreceptors in carotid sinus b. Photoreceptors: Rods and cones of the retina c. Chemoreceptors: Olfactory receptors, Taste receptors, Osmoreceptors , Carotid body O 2 receptors d. Thermoreceptors e. Pain receptors: Nociceptors
Receptive field It is an area of the body that, when stimulated, changes the firing rate of a sensory neuron. - Steps in sensory transduction a. Stimulus arrives at the sensory receptor. The stimulus may be a photon of light on the retina, a molecule of Na. Cl on the tongue and so on……. b. Ion channels are opened in the sensory receptor, allowing current to flow. Usually, the current is inward, which produces depolarization of the receptor. ■ The exception is in the photoreceptor, as light causes decreased inward current and hyperpolarization. c. The change in membrane potential produced by the stimulus is the receptor or generator potential. - If the receptor potential is depolarizing, it brings the membrane potential closer to threshold (firing level). -If the receptor potential is large enough, the membrane potential will exceed threshold, and an action potential will fire in the sensory neuron. Receptor potentials are graded depending on stimulus.
Adaptation of sensory receptors a. Slowly adapting, or tonic, receptors (muscle spindle; pressure) b. Rapidly adapting, or phasic, receptors (pacinian corpuscle for light touch) ■ show a decline in action potential frequency with time in response to a constant stimulus. - Sensory pathways from the sensory receptor to the cerebral cortex a. Sensory receptors: are activated by environmental stimuli. It may be specialized epithelial cells (e. g. , photoreceptors, taste receptors, auditory hair cells). Or it may be primary afferent neurons (e. g. , olfactory chemoreceptors). It transduce the stimulus into electrical energy (i. e. , receptor potential). b. 1 st order neurons: are the primary afferent neurons that receive the transduced signal and send the information to the CNS. Its Cell bodies are in dorsal root or spinal cord ganglia. c. 2 nd order neurons: are located in the spinal cord or brain stem. And receive information from one or more primary afferent neurons in relay nuclei and transmit it to the thalamus.
■Axons of second-order neurons may cross the midline in a relay nucleus in the spinal cord before they ascend to the thalamus. Therefore, sensory information originating on one side of the body ascends to the contralateral thalamus. d. 3 rd order neurons: are located in the relay nuclei of the thalamus. From there, encoded sensory information ascends to the cerebral cortex. e. 4 th order neurons: are located in the appropriate sensory area of the cerebral cortex. The information received results in a conscious perception of the stimulus. Motor Systems A. Motor unit -Consists of a single moto-neuron and the muscle fibers that it innervates. For fine control (e. g. , muscles of the eye), a single moto-neuron innervates only a few muscle fibers. For larger movements (e. g. , postural muscles), a single moto-neuron may innervate thousands of muscle fibers. ■ The motoneuron pool is the group of motoneurons that innervates fibers within the same muscle. ■ The force of muscle contraction is graded by recruitment of additional motor units (size principle). The size principle states that as additional motor units are recruited, more motoneurons are involved and more tension is generated.
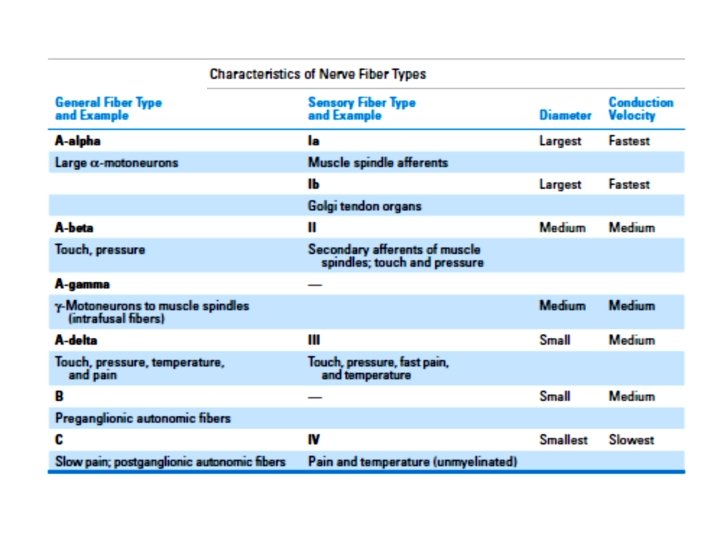
B. Muscle receptors 1. Types of muscle sensors : a. Muscle spindles (groups Ia and II afferents) are arranged in parallel with extrafusal fibers. They detect both static and dynamic changes in muscle length. b. Golgi tendon organs (group Ib afferents) are arranged in series with extrafusal muscle fibers. They detect muscle tension. c. Pacinian corpuscles (group II afferents) are distributed throughout muscle. They detect vibration. d. Free nerve endings (groups III and IV afferents) detect noxious stimuli. 2. Types of muscle fibers: a. Extrafusal fibers: make up the bulk of muscle. It is innervated by a-motoneurons. and provide the force for muscle contraction. b. Intrafusal fibers: are smaller than extrafusal muscle fibers. And are innervated by ɤ -motoneurons. are encapsulated in sheaths to form muscle spindles. run in parallel with extrafusal fibers, but not for the entire length of the muscle. They are too small to generate significant force.
Muscle Spindles are distributed throughout muscl, consist of small, encapsulated intrafusal fibers connected in parallel with large (force generating) extrafusal fibers. a. Types of intrafusal fibers in muscle spindles (1) Nuclear bag fibers -detect the rate of change in muscle length (fast, dynamic changes). - are innervated by group Ia afferents. - have nuclei collected in a central “bag” region. (2) Nuclear chain fibers -detect static changes in muscle length. are innervated by group II afferents. are more numerous than nuclear bag fibers. have nuclei arranged in rows.
b. How the muscle spindle works - Muscle spindle reflexes oppose (correct for) increases in muscle length (stretch). (1) Information about muscle length is received by group Ia (velocity) and group II (static) afferent fibers. (2) When a muscle is stretched (lengthened), the muscle spindle is also stretched, stimulating group Ia and group II afferent fibers. (3) Stimulation of group Ia afferents stimulates α-motoneurons in the spinal cord. This stimulation in turn causes contraction and shortening of the muscle. Thus, the original stretch is opposed and muscle length is maintained. c. Function of ɤ-motoneurons : innervate intrafusal muscle fibers. adjust the sensitivity of the muscle spindle so that it will respond appropriately during muscle contraction. α-Motoneurons and ɤmotoneurons are coactivated so that muscle spindles remain sensitive to changes in muscle length during contraction.
C. Muscle reflexes 1. Stretch (myotatic) reflex—knee jerk It is monosynaptic. a. Muscle is stretched, and the stretching stimulates group Ia afferent fibers. b. Group Ia afferents synapse directly on a-motoneurons in the spinal cord. The pool of α-motoneurons that is activated innervates the homonymous muscle. c. Stimulation of α-motoneurons causes contraction in the muscle that was stretched. As the muscle contracts, it shortens, decreasing the stretch on the muscle spindle and returning it to its original length. d. At the same time, synergistic muscles are activated antagonistic muscles are inhibited. e. Example of the knee-jerk reflex. Tapping on the patellar tendon causes the quadriceps to stretch. Stretch of the quadriceps stimulates group Ia afferent fibers, which activate α-motoneurons that make the quadriceps contract. Contraction of the quadriceps forces the lower leg to extend. ■ Increases in ɤ-motoneuron activity increase the sensitivity of the muscle spindle and therefore exaggerate the knee-jerk reflex. 2. Golgi tendon reflex (inverse myotatic) It is disynaptic. It is the opposite, or inverse, of the stretch reflex.
a. Active muscle contraction stimulates the Golgi tendon organs and group lb afferent fibers. b. The group Ib afferents stimulate inhibitory interneurons in the spinal cord. These interneurons inhibit a-motoneurons and cause relaxation of the muscle that was originally contracted. c. At the same time, antagonistic muscles are excited. d. Clasp-knife reflex, an exaggerated form of the Golgi tendon reflex, can occur with disease of the corticospinal tracts (hypertonicity or spasticity). ■ For example, if the arm is hypertonic, the increased sensitivity of the muscle spindles in the extensor muscles (triceps) causes resistance to flexion of the arm. Eventually, tension in the triceps increases to the point at which it activates the Golgi tendon reflex, causing the triceps to relax and the arm to flex closed like a jackknife.
3. Flexor withdrawal reflex (polysynaptic). It results in flexion on the ipsilateral side and extension on the contralateral side. Somatosensory and pain afferent fibers elicit withdrawal of the stimulated body part from the noxious stimulus. a. Pain (e. g. , touching a hot stove) stimulates the flexor reflex afferents of groups II, III, and IV. b. The afferent fibers synapse polysynaptically (via interneurons) onto motoneurons in the spinal cord. c. On the ipsilateral side of the pain stimulus, flexors are stimulated (they contract) and extensors are inhibited (they relax), and the arm is jerked away from the stove. On the contralateral side, flexors are inhibited and extensors are stimulated (crossed extension reflex) to maintain balance. d. As a result of persistent neural activity in the polysynaptic circuits, an after discharge occurs. The after discharge prevents the muscle from relaxing for some time.
Spinal organization of motor systems 1. Convergence: occurs when a single α-motoneuron receives its input from many muscle spindle group Ia afferents in the homonymous muscle. It produces spatial summation because although a single input would not bring the muscle to threshold, multiple inputs will. Also can produce temporal summation when inputs arrive in rapid succession. 2. Divergence: occurs when the muscle spindle group Ia afferent fibers project to all of the α-motoneurons that innervate the homonymous muscle. 3. Recurrent inhibition (Renshaw cells) ■ Renshaw cells are inhibitory cells in the ventral horn of the spinal cord. ■They receive input from collateral axons of motoneurons and, when stimulated, negatively feedback (inhibit) on the motoneuron.
Thank You