4 Proteini 4 1 Osobine proteina 4 2

4. Proteini § § § 4. 1. Osobine proteina 4. 2. Strukture proteina 4. 3. Klasifikaciija proteina
 od 100 do")
Šta su proteini ? § Proteini - su polimeri (makromolekuli, makropolipeptidi) od 100 do približno 1. 000 aminokiselinskih ostataka, što odgovara relativnoj molekulskoj masi (Mr) od 10 do 200 k. Da. § Nastaju povezivanjem ostataka aminokiselina peptidnom vezom (-CONH-) u obično duge polipeptidne lance (slika 4 -1):

§ Ime protein dao je Berzelius, a prvi put u literaturi se pominje daleke 1840. godine u udžbeniku Muldera. Ime se izvodi iz grčke reči proteos (prvi, primarni, najvažniji), a često se upotrebljava i izraz belančevine, pošto su proteini sastavni delovi belanceta jajeta. § Kod svih živih sistema pa time i biljaka proteini imaju dve esencijalne i različite uloge: oni predstavljaju strukturni materijal i pokazuju karakterističnu aktivnost i funkciju pa su samim tim i nosioci osnovnih životnih funkcija u ćeliji. Svi životni procesi ćelije se odvijaju u strukturama proteina ili uz njihovo učešće. Oni su i katalizatori brojnih biohemijskih procesa u ćeliji. Može se reći da ne poznajemo život bez proteina.

§ Proteini biljaka - se razlikuju međusobno u broju i redosledu aminokiselinskih ostataka, polipeptidnih lanaca, molekulskoj masi itd. U biljkama se nalazi u proseku oko 30% organske materije od čega na proteine kod pojedinih biljnih vrsta otpada u proseku 1520%. Proteini se nalaze ili slobodni u citoplazmi ili su asosovani sa drugim makromolekulima u ćelijskim organelama.

4. 1. Osobine proteina § Biološka funkcija proteina je u tesnoj vezi sa njihovim fizičko-hemijskim osobinama (Mr, polarnosti itd). § Mr proteina su veoma različite, a njihove brojne vrednosti su uslovljene metodom odredjivanja. Hordein iz ječma ima Mr 2. 700, edestin iz konoplje 31. 000, a ureaza iz soje 48. 000.

§ Proteini disosuju kao katjoni ili anjoni i mogu se smatrati polielektrolitima sa visoko izraženim amfolitnim svojstima (slika 4 -1). U zavisnosti od p. H rastvora reaguju kao kiseline ili baze. COOH NH COO NH COO NH + 3 - 2 - + 3 Katjon (p. H ≤ 7) Anjon (p. H ≥ 7) Dipolarni jon (p. H=7)

§ Proteini sa vodom grade koloidne rastvore. Stabilnost ovih koloida je različita i uslovljena je afinitetom prema vodi. Rastvori proteina u biljci imaju funkciju pufera velikog kapaciteta, a stabilnost u rastvoru je uslovljena njihovim afinitetom prema vodi odn. prisustvom hidrofilnih i hidrofobnih grupa u molekulu. U tabeli 4 -1 dati su neki primeri ovih grupa.

Tabela 4 -1. Hidrofilne i hidrofobne grupe u molekulu proteina: Hidrofilne grupe proteina Hidrofobne grupe proteina Karboksilna (-COOH) Metil (-CH 3) Karbonilna ( C=O) Metilenska (-CH 2 -) Hidroksilna (-OH) Etil (-C 2 H 5) Aldehidna (-CHO) Propil (-C 3 H 8) Amino (-NH 2) Fenil (-C 6 H 5) Imino ( NH) Izopropenska (-C 5 H 8) Amidna (-CONH 2) Sulfhidrilna (-SH)
 proteini")
§ Proteini mogu biti ekstremno nerastvorljivi ili dobro rastvorljivi. Pri većim temperaturama (kuvanjem) proteini denaturišu, pri čemu se povećava njihova nerastvorljivost u vodi. Zbog relativno velike molekulske mase proteini mogu graditi sole i gele. § Pomoću polarnih grupa proteini mogu vezati i do 30 g vode na 100 g proteina (vezana voda se na niskim temperaturama ne može smrznuti). Kada se vezuju sa drugim jedinjenjima grade komplekse koji su od velikog značaja za ćelijske i međućelijske strukture. Može se reći da je individualnost svakog organizma uslovljena proteinima koji ga izgrađuju.

4. 2. Strukture proteina § S obzirom na to da su proteini makromolekuli, pojam strukture je kompleksniji u poređenju s prostim organskim molekulima i obuhvata hijerarhijsku strukturnu organizaciju u četiri nivoa: § primarna, § sekundarna, § tercijarna § kvaternerna struktura § Prostorna građa molekula proteina se određuje fizičkim ili fizičkohemijskim metodama, najčešće rendgeno-strukturnom analizom. Opisivanje ovih struktura obuhvata dva aspekta: § opisivanje konformacije molekula i § sila koje uslovljavaju pojedine strukture.

§ Sastav i konformacija molekula proteina određuje njihovu biološku funkciju. § Niz specifičnih osobina proteina (enzimsko delovanje, hormonska aktivnosti i sl. ) zavise od redosleda (sekvence) aminokiselina u polipeptidnin nizovima i od njihovog prostornog rasporeda. U nekim slučajevima specifična aktivnost proteina ne zavisi od sekvence aminokiselina u čitavom polipeptidnom nizu, već samo od redosleda u nekim delovima niza koji se može javiti i na više mesta u molekulu (tzv. reaktivni ili aktivni centri).
")
1. Primarna struktura § jednodimenzionalna struktura (predstavlja prvi stepen u specifičnoj trodimenzionalnoj strukturi proteina) označava vrstu i redosled (sekvencu) aminokiselina i položaj S S veza, a po novijem i udeo pojedinih aminokiselina u polipeptidnom lancu. § Peptid H 2 N-Leu-Gly-Thr-Val-Arg-Asp-His-COOH se razlikuje u primarnoj strukturi od peptida H 2 NVal-His-Asp-Leu-Gly-Arg-Thr-COOH, čak i ako oba imaju isti broj i iste vrste aminokiselina, ali se razlikuju u njihovom redosledu. Primarna struktura je stabilizovana peptidnim vezama.

§ Osim toga konfiguracija i stabilnost proteina izgrađenog iz više peptidnih lanaca uslovljena je i sekundarnim (bočnim) vezama. Ove veze se nazivaju sekundarnim jer osnovnu, primarnu vezu predstavlja peptidna veza, kojom su aminokiseline povezane u duge polipeptidne nizove. Polipeptidni nizovi poseduju bočne funkcionalne grupe koje reaguju sa bočnim funkcionalnim grupama drugog (ili ako je savijen istog) niza, gradeći sekundarne veze (vodonična, disulfidna, jonska, hidrofobna itd. ) (slika 4 -2).

§ Slika 4 -2. § Sekundarne veze kod proteina. Peptidni lanci su prikazani linijama: 1 - vodonična veza između peptidnih veza, 2 - disulfidna veza između dva cisteinska ostatka, 3 - jonsko privlačenje između bočnih grupa Asp i Lys, 4 - hidrofobna veza između valinskog i izoleucinskog ostatka, 5 – estarska veza.

§ Sekvenca aminokiselina je genetski predodređena. Proteini koji imaju slične sekvence nazivaju se homologi proteini. Oni imaju često i sličnu funkciju. Homologija proteina se može objasniti činjenicom da su se oni u toku evolicije sintetizovali iz istog prekursora. Poređenje sekvence među homologim proteinima može da otkrije genetsku vezu između vrsta.
 (slika")
§ Prvi peptid kome je određena primarna struktura bio je insulin (Sanger, 1953) (slika 4 -3), a prvi protein, ribonukleaza iz pankreasa (Hirs, Stein i Moor, 1959).

Slika 4 -3. Hipotetično filogenetsko stablo citohroma c po L. Harperu I P. Weissu.
 polipeptidnog lanca u prostoru. § Način")
2. Sekundarna struktura § predstavlja način uvijanja (nabiranja) polipeptidnog lanca u prostoru. § Način nabiranja je uslovljen sekvencom aminokiselinskih ostataka, kao i “silama” koje povezuju različite odsečke polipeptidnog lanca. § Drugim rečima ona predstavlja način nabiranja polipeptidnog lanca u prostoru, kao i uvrtanje amidnih ravni oko veze koja spaja -C atome proteinskih lanaca. Polipeptidna kičma predstavlja povezanu sekvencu čvrstih planarnih peptidnih grupa (slika 4 -4), tako da svaki aminokiselinski ostatak u polipeptidnom nizu ima dva stepena slobode, tj. moguće su rotacije oko dve veze: C -C’ ( ) i C -N ( ) (slika 4 -4). Ovi uglovi se nazivaju rotacioni ili torzioni uglovi.

Slika 4 -4. Polipeptidni niz u svojoj maksimalnoj istegnutoj konformaciji. Slika 4 -5. Segment polipeptidnog niza sa obeleženim torzionim (rotacionim) uglovima oko veza C -N ( ) i C -C ( ).

§ Način nabiranja polipeptidnog lanca je uslovljen sekvencom aminokiselinskih ostataka kao i “silama” koje povezuju različite odsečke polipeptidnog lanca (vodonične, jonske, hidrofobne veze i Van der Waalsove sile).

§ Proteini se sintetizuju sekvencijalnom adicijom aktiviranih aminokiselina u rastući polipeptidni lanac i umotavaju u strukturu nativnog proteina. Način umotavanja uslovljen je primarnom strukturom koja omogućuje stvaranje određenih hemijskih veza. Umotavanje polipeptidnog lanca se realizuje u tri tipa sekundarnih struktura i to: § -heliks ( -uzvojnica), § -plisirana struktura (nabrana -konformacija) i § -obrt. § Delovi proteina koji se ne umotavaju ni u jedan oblik sekundarne strukture nazvani su “random coil” ili struktura slučajnog namotavanja klupčeta odn. neuređena struktura. Najrasprostranjeniji oblik sekundarne strukture je -uzvojnica koja je energijom najsiromašnija i najstabilnija konformacija proteina.

§ -heliks - je najpoznatiji oblik sekundarne strukture proteina koju je identifikovao Linus Pauling 1951 g. i ona uključuje strukturu u prostoru samo jednog polipeptidnog lanca. § “Kičma” lanca je aranžirana u heliks sa 3, 6 aminokiselinskih ostataka u svakom obrtu. U heliksu svaka peptidna imino grupa (-NH-) jedne peptidne veze je povezana vodoničnom vezom za peptidnu karbonilnu grupu ( C=O) neke susedne peptidne veze. Vodonične veze se stvaraju između pojedinih navoja što daje ovoj strukturi posebnu stabilnost (slika 4 -7).

§ Slika 4 -7. § Intravodonične veze i druge sekundarne veze daju polipeptidnom lancu konfiguraciju -spirale. § Vodonične se veze stvaraju između pojedinih navoja (peptidne veze se nalaze jedna iznad druge) i to daje -spirali posebnu stabilnost. -Heliks ima 3, 6 ostataka aminokiselina po okretu, a rastojanje između dva okreta je 0, 54 nm. Poluprečnik spirale je 0, 25 nm, a ugao između ravni u kojima se nalaze dve susedne peptidne veze je 1080. Vodonične veze nastaju između amidnih Hatoma i O-atoma karbonilnih grupa, koji su u nizu udaljeni 4 peptidne veze.
 u kojoj su vodonične")
Slika 4 -8. Loptasto-štapićast model desne - heliks spirale: (a) u kojoj su vodonične veze između N H grupe iz n-tog aminokiselinskog ostatka i C=O iz n+4 -tog aminokiselinskog ostatka obeležene isprekidanim linijama i (b) model strukture proteina sa naznačenim heliksnim regionima

§ Istraživanja L. Paulinga su takodje pokazala da proteini pored -heliksa mogu imati i -strukturu koja je dobila čitav niz imena kao npr. struktura presavijenih površina, nabrana, plisirana struktura itd. § -Struktura - se obrazuje kada se dva ili više polipeptidna lanca ili segmenta jednog istog lanca povežu uzduž jedan sa drugim vodoničnim vezama. Vodonične veze su normalne u odnosu na kičmu polipeptidnog lanca. Polipeptidni lanac u -nizu je skoro sasvim razvučen (cik-cak konformacija)

§ Razmak između -nabrana struktura aminokiselina duž ose je 0, 35 nm. Sve peptidne grupe učestvuju u formiranju vodoničnih veza i tako održavaju strukturu stabilnom. Bočne grupe aminokiselinskih ostataka leže između ili ispod ravni nabrane površine.
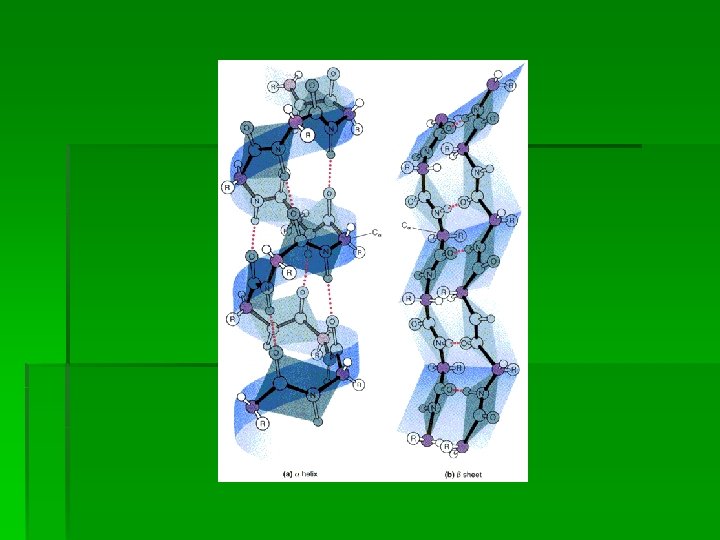

§ -nabrana struktura može biti: antiparalelna - u kojoj su susedni, vodonično vezani, polipeptidni nizovi suprostavljenih smerova i paralelna - u kojoj su susedni, vodonično vezani, polipeptidni nizovi istog smera Slika 4 -9. -Nabrana struktura proteina (a) Vodonične veze su prikazane isprekidanom linijom, (b) Vodonične veze u antiparalelnoj i (c) Vodonične veze u paralelnoj -pločici sastavljenoj iz tri niza

Slika 4 -10. Trodimenzionalni oblik -strukture. Slika 4 -11. Povezivanje susednih nizova u -pločici: (a) povezivanje susednih antiparalelnih peptidnih nizova tipa ukosnice; (b) desna petlja medju paralelnim nizovima i (c) leva petlja medju paralelnim nizovima.

§ Pored -heliksa i -pločice u globularnom proteinu neki deo strukture se može nalaziti i u obliku -obrta ( zavijutka). § -zavijutak ( -obrt ) - oko polovine polipeptidnog niza u prosečnom globularnom proteinu uvijeno je u obliku -heliksa i -pločice, dok se preostali deo većinom nalazi u obliku -zavijutka. § -zavijutak - se javlja se kao tip 1, tip 2 i tip 3. U njemu su najčešće zastupljene aminokiseline glicin, prolin, asparagin i serin. Ovaj tip prostorne strukture proteina se javlja u globularnim proteinima.

§ Učešće pojedinih AK u sekundarnim strukturama:

§ Omega petlja - Skoro svi globularni proteini koji sadrže više od 60 aminokiselinskih ostataka sadrže jednu ili više petlji koje se sastoje iz 6 do 16 aminokiselinskih ostataka u kojima su zastupljene prostorne strukture tipa -heliksa i pločice. Takva petlja koja često sadrži i -zavijutke, se zbog svog oblika naziva i omega petlja. Omega petlja
.")
Kombinacijom -heliksa i -pločice mogu nastati različiti oblici supersekundarne strukture proteina (slika 4 -14). Šematski dijagrami supersekundarnih struktura proteina. Strelice pokazuju smer polipeptidnog lanca. (a) jedinica, (b) jedinica, (c) -obrt i (d) grčki ključ.
 -motiv, b) ukosnica, c) motiv,")
Slika 4 -14 b. Šematski prikaz supersekundarnih struktura. a) -motiv, b) ukosnica, c) motiv, d) grčki ključ. Strelice pokazuju N-C pravac polipeptidnog lanca § Asocijacijom sekundarnih strukturnih elemenata proteina ( spirale i -nabrane strukture) pomoću interakcija bočnih lanaca mogu nastati različiti oblici supersekundarne strukture proteina (motiv), slika 4 -7.
 predstavlja viši oblik organizacije proteina u prostoru u")
3. Tercijarna struktura § (trodimenzionalna struktura) predstavlja viši oblik organizacije proteina u prostoru u kojoj se nabiraju segmenti heliksa, plisirane strukture i -obrta peptidnog lanca u trodimenzionalnu strukturu nativnog proteina ili proteinske subjedinice stvarajući globule ili fibrile.
 sekundarnih struktura i")
§ Može se reći da tercijarna struktura predstavlja način organizacije (rasporeda) sekundarnih struktura i položaj bočnih ostataka aminokiselina u molekulu datog proteina. Kod proteina koji se sastoje iz samo jedne globule (jednog polipeptidnog lanca) to je najviši oblik strukture u prostoru. § Tercijarna struktura proteina zahteva potpuno uređen trodimenzionalni raspored svih atoma u molekulu proteina. Ova struktura stabilizovana je pored vodonične i drugim energetski još povoljnijim hemijskim vezama izmedu R ostataka aminokiselina u polipeptidnom lancu kao što su jonske, kovalentne, hidrofobne veze i disperzne sile (slika 4 -2).

§ Tercijarna struktura proteina se određuje rendgenostrukturnom analizom i ustanovljena je za veći broj proteina. Promena tercijarne strukture uslovljava promenu bioloških (npr. enzimsko ili hormonsko delovanje) i fizičkih osobina proteina. § Proteini se klasifikuju na osnovu tercijarne strukture na globularne, koji mogu biti loptasti ili elipsoidni, i fibrilarne koji mogu biti jednostruki, dvostruki ili višestruki (slika 416). Tercijarna struktura proteina: (a) globularna, (b) fibrilarna
 proteini su strukturni proteini kod kojih su odnosi osa veći od")
§ Fibrilarni (fibrilni) proteini su strukturni proteini kod kojih su odnosi osa veći od 10. Nastaju preplitanjem nekoliko polipeptidnih lanaca. Fibrilarni proteini su jako izduženi (sl. 4 -16 b) i pretežno se javljaju u obliku sekundarne strukture. Oni grade vlakna, od kojih su neka ugrađena u matrice od organskog ili neorganskog materijala, što dodatno doprinosi čvrstoći bioloških struktura u kojima se ovi molekuli nalaze. Fibrilni proteini, imaju strukturnu i zaštitnu ulogu, a predstavljaju ključne komponente u biološkim strukturama. Fibrilarni proteini su nerastvorni, i ne mogu se dobiti u kristalnom stanju, što znatno otežava njihovo izolovanje i ispitivanje
 proteini su oni proteini kod kojih su odnosi njihovih osa (dužina")
§ Globularni (sferoidni) proteini su oni proteini kod kojih su odnosi njihovih osa (dužina : širina) manji od 10: 1 (slika 4 -16 a). Sastoje se iz peptidnih lanaca koji su izuvijani i namotani na kompaktan način. Molekuli su manje ili više sfernog oblika, slično elipsoidu ili lopti. Međusobni odnos njihovih osa nije veći od 3: 1 ili 4: 1. Globularni proteini su enzimi i drugi brojni biološki aktivni proteini. Kod globularnih proteina hidrofobne grupe su orijentisane unutar globule, a polarne su na površini molekula što omogućava rastvorljivost ovih proteina u vodi. Rendgenskom analizom određena je tercijarna struktura manjeg broja proteina npr.

§ Prema vrsti domena koje sadrže proteini tercijarne strukture se mogu klasifikovati u 4 grupe: proteine u kojima preovladava struktura -uzvojnice, proteine u kojima preovladava -nabrana struktura, proteine u kojima se -uzvojnica nalazi u jednom delu strukture, a u drugom -nabrana struktura i proteine u kojima se naizmenično smenjuju -heliks i -pločica.
.")
Slika 4 -17. Tercijarna struktura gliceraldehid 3‑fosfat dehidrogenaze (pojednostavljen prikaz supersekundarne strukture enzima).

4. Kvaternerna struktura § je finalni oblik strukture proteina nastala interakcijom različitih polipeptidnih lanaca. § veliki proteini (npr. globularni proteini mase veće od 100 k. Da), skoro po pravilu se javljaju u obliku agregata koji se sastoje iz subjedinica asosovanih na geometrijski određen način. Naziv kvaternerna struktura za proteine koji se sastoje iz nekovalentno vezanih subjedinica, uveo je Bernal, 1958 godine. Mnogi rastvorljivi proteini, a i neki nerastvorljivi, su izgrađeni iz dva ili više (dimer, trimer ili tetramer) polipeptidnih lanaca (subjedinica) koji mogu biti isti ili različiti. Ove subjedinice se mogu izolovati iz koncentrovanog rastvora soli ili pod blagim uslovima denaturacije. U tabeli 4 -2 dat je pregled nekih proteina koji su izgrađeni iz dve i više subjedinica.

Tabela 4 -2. Proteini sa dve i više subjedinica. Vrsta proteina Mr proteina Broj subjedinica Nitrat-reduktaza 197. 000 2 Aldolaza 142. 000 3 Heksokinaza 96. 000 4 Gliceraldehid-3 -fosfat dehidrogenaza 140. 000 4 Ureaza 483. 000 6 Glutamat-dehidrogenaza 2. 000 8 Mozaik virus duvana 40. 000 2130
 aspartat-amonijumliaze (aspartaza) § Uopšten oblik kvaternerne")
Slika 4 -15. Model kvaternarne strukture proteina (enzima) aspartat-amonijumliaze (aspartaza) § Uopšten oblik kvaternerne strukture proteina može se prikazati pomoću modela enzima aspartatamonijumliaze (EC 4. 3. 1. 1) u kojem je svaka podjedinica predstavljena posebnom loptom (slika 4 -15).

§ Dovoljno je da se samo jedna subjedinica izdvoji iz agregata kvaternerne strukture molekula pa da enzim izgubi svoju biološku funkciju. Aspartatamonijumliaza ili aspartaza (EC 4. 3. 1. 1) katalizuje reversnu konverziju aspartata u fumarat:
 sekundarna, (b) tercijarna, (c) kvaternerna")
Slika 4 -16. Strukture proteina: (a) sekundarna, (b) tercijarna, (c) kvaternerna
 § Najbolje")
Slika 4 -17. Kvaternerna struktura Hb i -subjedinice hemoglobina ( 1 2) § Najbolje je proučena kvaternerna struktura hemoglobina (Hb) kao model strukture globularnog proteina (slika 417).

Slika 4 -18. Denaturacija proteina

4. 3. Klasifikacija proteina § Proteini biljaka su osnovna hrana ljudi i životinja jer su jedini izvor esencijalnih aminokiselina. U ishrani ljudi biljni proteini su zastupljeni sa 80%, proteini životinja sa 15%, a riba sa 5%. Do danas je iz biljaka izolovan veliki broj proteina, te se javlja potreba za njihovom klasifikacijom. Zbog nepoznavanja primarne strukture većine proteina, nije se mogla uvesti hemijska klasifikacija te je pokušano da se biljni proteini klasifikuju prema: složenosti, subcelularnoj lokaciji, funkciji, elektroforetskoj pokretljivosti i dr.

§ Nijedna od navedenih klasifikacija nije kompletna i one se medjusobno mogu dopunjavati. Medjutim u praksi se najčešće koristi klasifikacija proteina prema složenosti, na osnovu koje se proteini mogu podeliti u dve grupe i to: proste (nekonjugovane ) proteine i složene (konjugovane) proteine Prosti (nekonjugovani) proteini - daju hidrolizom samo aminokiseline i dele se na globularne i fibrilarne proteine. Još je Osborn 1924. godine globularne proteine biljaka klasifikovao na osnovu njihove rastvorljivosti i osobina koja je još i danas u upotrebi u četiri grupe (tabela 4 -3).

Naziv proteina Albumini Rastvorljivost Sastav Rastvorni u destilovanoj vodi pri neutralnim ili slabo kiselim p. H, zagrevanjem lako koagulišu. Sadrže pretežno Glu, Lys, Leu (15%), a ne sadrže Gly. Nalaženje U biljkama se nalaze u količlini 0. 6 -2. 0% i to manje u zelenim delovima, a više u zrnu kao leukozin u pšenici, raži, ječmu i legumelin u grašku. To su tipični enzim-proteini. Globulini Nerastvorni u vodi, a rastvorni u razblaženim rastvorima soli, zagrevanjem teško koagulišu Sadrže pretežno Leu (4%) Val, Lys, Ser, Glu, a ne sadrže Gly. Veoma su rasprostranjeni u biljkama i to kao legumin u grašku, fazeolin u pasulju, konglutin u lupini, glicinin u soji i edestin u konoplji. Glutelini Nerastvorni u vodi, Ne sadrže Lys rastvorima soli i ali su Et. OH, a rastvorni u bogati u jakim kiselinama i Pro, Asp i bazama Glu. U biljkama se nalaze od 1 -3%: orizenin u pirinču, vicilin u grašku, glutenin u pšenici i kukuruzu. To su rezervni proteini semena. Prolamini Rastvorni su u 70% etanolu, ali ne i u absolutnom etanolu Sadrže najviše Pro i Glu. U zrnastim kulturama i to: avein u ovsu, gliadin u pšenici, hordenin u ječmu. To su rezervni proteini semena.
 proteini ili proteidi - sadrže pored proteinskog i neproteinski deo koji")
§ Složeni (konjugovani) proteini ili proteidi - sadrže pored proteinskog i neproteinski deo koji se naziva prostetičnom grupom. § Ona je obično vezana za proteinski deo molekula kovalentnim, heteropolarnim ili koordinativnim hemijskim vezama. Prostetična grupa može biti neki ugljeni hidrat, lipid, nukleinska kiselina, metal, pigment, fosfatni ostatak i dr. Prema prirodi prostetične grupe složeni proteini (konjugovani proteini ili proteidi) se mogu podeliti u nekoliko grupa i to: nukleoproteine, glikoproteine, lipoproteine metaloproteine i dr.

§ Nukleoproteini - su složeni proteini jedra, hromozoma, ribozoma i mitohondrija. Oni kao neproteinsku komponetu sadrže nukleinske kiseline. Sastoje se iz baznog proteina histona ili prolamina čije se relativne molekulske mase kreću od 10. 000 do 20. 000 pa i više. U biljkama histoni se javljaju u osnovna tri strukturna oblika i to kao: - histoni bogati Lys, - histoni sa manjim sadržajem Lys i - histoni bogati Arg. § Obično u svojoj strukturi histoni sadrže više stotina aminokiselinskih ostataka (najčešće od 100 - 250) i sa nukleinskim kiselinama grade jedinjenja tipa soli koja omotavaju i štite spiralu DNA.

Lipoproteini - sadrže lipid kao neproteinsku komponentu koji je veoma čvrsto vezan za ostatak proteina tako da se ova dela molekula veoma teško razdvajaju elektroforezom, ultracentrifugovanjem i taloženjem solima teških metala. Lipidna komponenta je najčešće lecitin, kefalin ili holesterol. Odnosi proteinskog dela prema lipidnom variraju od 20: 80 do 75: 25. Lipoproteini se najvećim delom nalaze u ćelijskoj membrani, mitohondrijama. . . Slika 4 -22. Fluidno-mozaični model membrane po Singeru.

§ Glikoproteini - sadrže ugljene hidrate od kojih najčešće oligosaharide kao prostetičnu grupu. Oni mogu biti sastojci ćelijskih membrana, enzimi (kao npr. peroksidaza), lektini (fitohemaglutin koji izaziva aglutinaciju eritrocita), hormoni, otrovni proteini (ricin iz semena ricinusa) komponente sluzi i dr. Mnogi glikoproteini transportuju supstance kroz membrane.

§ Metaloproteini - su konjugovani proteini sa metalima. U metaloproteinima metali su funkcionalni sastojci ovih molekula i obično nosioci njihovih katalitičkih i regulatornih funkcija. Značajniji metaloproteini koji sadrže gvožđe su citohromi, respiratorni pigmenti, enzimi (npr katalaza i dr) itd. Cink je prisutan u karboanhidrazama, a neki glikolitički enzimi i proteaze sadrže magnezijum. § Pored napred navedenih klasifikacija proteini se mogu klasifikovati i po osnovu njihove biološke funkcije na više grupa koje obuhvataju: enzime rezervne proteine (proteini semena), metaboličke proteine (proteini citoplazme), membranske proteine itd.

§ U novije vreme se proteini klasifikuju i prema subcelularnoj lokaciji. Njihov procentni udeo u nekim značajnim organelama je sledeći: 40 -60% u hloroplastima, 35 -40% u mitohondrijama i 1 -2% u jedru § Proteini se mogu klasifikovati i prema elektroforetskoj pokretljivosti. Elektroforeza je kretanje naelektrisanih molekula proteina u rastvoru pod uticajem jakog električnog polja koje je uslovljeno naelektrisanjem i molekulskom masom proteina.

§ Od biljnih proteina od posebnog su značaja proteini ratarskih kultura. Neki od njih mogu biti inhibitori proteolitičkih enzima sisara (kao npr. tripsin inhibitor soje, TIS) i tako smanjiti hranljivu vrednost biljnih proteina. Tabela 4 -5. Procentni udeo proteina u odnosu na suvu masu: Kultura Udeo proteina (%) Kukuruz Pšenica Ricinus Suncokret Soja 10 12 16 -24 15 -30 30 -40
 2%, ►kupus (Brassica oleraceae)")
§ Povrće - sadrži relativno malo proteina: ►krompir (Solanum tuberosum) 2%, ►kupus (Brassica oleraceae) 1. 5%, ►mrkva (Daucus carota) 1. 5% ►krastavac (Cucumis sativus) 0. 01%, ►paprika (Capsicum annum) 1. 2 -1. 6%.

§ Kvalitet proteina biljaka - se ceni prema količini esencijalnih aminokiselina i njihovoj svarljivosti. Oni mogu biti “nepotpuna hrana” ukoliko su deficitarni u nekim aminokiselinama. Tako npr. proteini trava su deficitarni u svim aminokiselinama, zrna kukuruza u Lys, graška u Trp i Met, pasulja u Lys i sl. § Proteini pšenice sadrže polovinu potrebnog Lys za čoveka, a proteini krompira i graška trećinu Met i Cys. Procentni udeo esencijalnih aminokiselina u proteinima suncokreta, lucerke i spanaća dat je u tabeli 4 -6.
 kukuruza")
Tabela 4 -6. Udeo esencijalnih aminokiselina u proteinima ratarskih kultura. Aminokiselina Zein (%) kukuruza Valin Leucin Izoleucin Fenilalanin Metionin Triptofan Lizin 1. 9 2. 9 7. 6 2. 4 1. 8 - Gliadin pšenice Suncokret Lucerka Spanać 3. 0 6. 0 2. 5 2. 3 1. 4 8. 2 5. 1 6. 4 4. 3 4. 5 1. 4 3. 6 5. 0 6. 3 3. 7 3. 3 1. 0 7. 9 6. 1 9. 5 4. 9 6. 1 2. 1 7. 3

§ Proteini biljaka - biće i u budućnosti glavna hrana za čoveka i životinje i pored toga što se planiraju novi izvori proteina – sintetički proteini kao i proteini iz parafina nafte, planktona mora i dr. Osim za ishranu proteini su značajni za primenu kao antimetaboliti (toksični proteini) i kao zaštitna sredstva protiv bolesti (jer inaktiviraju viruse, bakterije i sl. agense). Zbog navedenog se očekuje da se u budućnosti proizvodnja proteina biljaka udvostruči.
- Slides: 62